Передача РНК (тРНК) – определение, типы и функции
Определение тРНК
Трансферные РНК или тРНК представляют собой молекулы, которые действуют как временные носители аминокислоты в результате чего соответствующие аминокислоты рибосома на основе мессенджера РНК (мРНК ) нуклеотид последовательность. Таким образом, они действуют как посредники между нуклеотидной и аминокислотной последовательностями.
тРНК представляют собой рибонуклеиновые кислоты и поэтому способны образовывать водородные связи с мРНК. Кроме того, они также могут образовывать сложноэфирные связи с аминокислотами и, следовательно, могут физически объединять мРНК и аминокислоты в процессе перевод, Они спариваются с мРНК комплементарным и антипараллельным образом, и каждая тРНК может базовая пара с натяжкой из трех нуклеотидов на мРНК. Эти наборы из трех нуклеотидов на мРНК называются кодонами, а соответствующая последовательность на тРНК называется антикодоновую, Спаривание оснований между кодоном и антикодоном вносит специфичность в процесс трансляции. На одном конце тРНК соответствующая аминокислота присоединена к ее 3 ’ гидроксильная группа основанный на антикодоне и рибосоме катализирует образование пептидная связь между этой аминокислотой и удлиненной полипептидной цепью.
Структура и функции тРНК
Трансферные РНК кодируются рядом генов и обычно представляют собой короткие молекулы длиной от 70 до 90 нуклеотидов (5 нм). Двумя наиболее важными частями тРНК являются ее антикодон и концевая 3′-гидроксильная группа, которая может образовывать сложноэфирную связь с аминокислотой. Тем не менее, существуют и другие аспекты структуры тРНК, такие как D-плечо и T-плечо, которые способствуют его высокому уровню специфичности и эффективности. Только 1 из 10000 аминокислот неправильно присоединены к тРНК, что является замечательным числом, учитывая химическое сходство между многими аминокислотами.
Трансферные РНК имеют сахарно-фосфатный остов, как и все другие клеточные нуклеиновые кислоты, и ориентация рибозного сахара вызывает направленность в молекула, Один конец РНК имеет реактивный фосфатная группа присоединен к пятому атому углерода рибозы, тогда как другой конец имеет свободную гидроксильную группу на третьем атоме углерода. Это приводит к 5 ‘и 3’ концам РНК, так как все другие фосфатные и гидроксильные группы участвуют в фосфодиэфирных связях внутри нуклеиновая кислота.


D-рычаг состоит из двухцепочечной области ствола, образованной внутренним спариванием оснований, а также петлевой структурой неспаренных нуклеотидов. D-дуга является сильно изменчивой областью и играет важную роль в стабилизации РНК. третичная структура а также влияет на кинетику и точность трансляции на рибосоме.
Другой структурой, которая влияет на роль тРНК в трансляции, является Т-образный рычаг. Как и у D-плеча, он содержит участок нуклеотидов, пары оснований которых соединены друг с другом, и петлю, которая является одноцепочечной. Парный регион называется «стебель» и в основном содержит 5 пар оснований. Цикл содержит модифицированные основания и также называется плечом TΨC, чтобы указать присутствие остатков тимидина, псевдоуридина и цитидина (модифицированных оснований). Молекулы тРНК необычны тем, что содержат большое количество модифицированных оснований, а также содержат тимидин, обычно наблюдаемый только в ДНК. Т-образный рычаг участвует во взаимодействии тРНК с рибосомой.
Наконец, вариабельное плечо, содержащее менее 20 нуклеотидов, расположено между антикодонной петлей и Т-образным плечом. Он играет роль в распознавании тРНК ААТС, но может отсутствовать в некоторых вид.
Считается, что вторичная структура тРНК, содержащая акцепторную область, D- и T-плечи и петлю антикодона, напоминает клеверный лист. После того как РНК складывается в свою третичную структуру, она становится L-образной, с акцепторным стержнем и Т-образным плечом, образующим расширенную спираль, а антикодонная петля и D-плечо аналогично образуют еще одну расширенную спираль. Эти две спирали располагаются перпендикулярно друг другу таким образом, что D-рычаг и Т-образный рычаг находятся в непосредственной близости, в то время как петля антикодона и плечо акцептора расположены на противоположных концах молекулы.

На этом изображении 3’’-область CCA выделена желтым цветом, плечо акцептора – фиолетовым, переменная петля – оранжевым, D-плечо красным, T-плечо зеленым и антикодонная петля синим.
Типы тРНК
ТРНК может быть классифицирована на основе аминокислоты, которую она несет, давая в результате 20 различных тРНК. Альтернативно, они также могут быть сгруппированы на основе их антикодона. Существует 64 возможных кодона, возникающих из комбинации четырех нуклеотидов. Из них 3 являются стоп кодоны это сигнал об окончании перевода. Это приводит к ситуации, когда одна аминокислота представлена множественными кодонами и AATS, а также тРНК должны учитывать эту избыточность. Однако очень немногие виды имеют ровно 61 тРНК, что ставит вопрос о том, как каждый кодон распознается конкретной тРНК. У многих видов это число значительно превышает 61, и разные тРНК, несущие один и тот же антикодон, могут демонстрировать различную эффективность в трансляции, добавляя слой регуляции в процесс синтеза белка.
тРНК взаимодействуют с кодонами мРНК через их антикодонную петлю. Спаривание оснований между кодоном и антикодоном обеспечивает специфичность при трансляции. Однако первое основание антикодона, которое соединяется с «колебанием» или третьей позицией в кодоне, часто модифицируется, чтобы позволить тРНК образовывать водородную связь с тремя, а не с одним основанием. Таким образом, одна тРНК имеет возможность распознавания и спаривания оснований с тремя кодонами, которые кодируют одну и ту же аминокислоту. Есть 20 ААТС, по одному на каждую аминокислоту. Эта группа ферментов может распознавать все антикодоны, представляющие определенную аминокислоту, и, следовательно, выступать в качестве второго рычага механизма, который обрабатывает генетический код избыточность.
Наконец, эти молекулы также можно разделить на три категории: те, которые несут канонические аминокислоты, присоединенные к правильной тРНК, те, которые неправильно присоединены, и те, которые несут модифицированные аминокислоты, такие как селеноцистеин, для неканонического удлинения.
Посттранскрипционная модификация тРНК
В геноме человека насчитывается около 500 генов, кодирующих тРНК, и 300 ген фрагменты, связанные с этими РНК. Эти гены транскрибируются РНК-полимераза III и транскрипт подвергается обширной модификации, особенно у эукариот. Интроны сращены, интрон –экзон На границу действуют эндонуклеазы, 5′- и 3′-концы РНК обрабатываются, и ферменты добавляют концевые остатки CCA к 3′-концу тРНК. Остатки CCA могут стать аминоацилированными в самом ядре, и эта заряженная тРНК может затем экспортироваться из ядра.
Кроме того, многие основания на тРНК также модифицируются, особенно метилированием (добавление метильной группы) и дезамидированием (удаление амидной группы). В частности, первая база антикодона, которая спаривается с положением «колебания» на кодоне, модифицирована, чтобы допустить необычные типы спаривания оснований. Аденин может быть модифицирован с образованием инозина, который расширяет возможности соединения, включая урацил, цитозин и аденин. Псевдоуридин является еще одним распространенным модифицированным основанием, полученным из остатков уридина посредством ферментативно-опосредованной изомеризации. Говорят, что он играет роль в структурной целостности молекулы тРНК, участвует в придании жесткости близлежащему сахарофосфатному остову, а также влияет на укладку основания проксимальных областей. Лизидин является необычным основанием, образованным, когда аминокислота лизина присоединена к остатку цитидина. Лизидин специфически связывается с аденозином, свойство, которое используется тРНК изолейцина для обеспечения специфичности трансляции.
AATS присоединяют подходящую аминокислоту к молекулам тРНК на основе их антикодона. Эти ферменты содержат сайты связывания для аминокислоты, тРНК, а также АТФ и Hydrolyze ATP к AMP и присоедините аминокислоту к рибозному сахару последнего нуклеотида на тРНК. Теперь тРНК считается «заряженной» и может участвовать в реакциях синтеза белка на рибосоме. Эта реакция часто происходит в цитоплазма Хотя это также наблюдалось в ядре.

Зрелая тРНК затем связывает специфические экспортные факторы, которые экспортируют ее из ядра, используя систему RanGTP. Рука акцептора и T-рука играют важную роль в этом процессе, и существует обширное взаимодействие между факторами экспорта и молекулой РНК, что позволяет только полностью обработанным, полным тРНК перемещаться в цитоплазму.
Взаимодействие тРНК с рибосомой
Рибосома содержит три важных области – P (пептидильный) сайт, содержащий растущий полипептид, A (акцепторный) сайт, который получает новую заряженную тРНК, и E (выходной) сайт, через который деацилированная тРНК покидает рибосому. Эти сайты охватывают обе субъединицы рибосомы и обозначаются как сайты P / P или A / A, причем первая буква относится к сайту на меньшей субъединице. Например, сайт P / P связывается с тРНК, закрепляющей полипептидную цепь, в то время как сайт A / A привязывает входящую заряженную тРНК. Пептидил-тРНК в P / P-сайте переносит растущий полипептид в тРНК в A / A-сайте и подвергается деацилированию. Чтобы продолжить процесс трансляции, рибосома продвигается вперед на один кодон, заставляя тРНК в P / P-сайте сместиться в переходную P / E-конфигурацию и затем в E / E-сайт, прежде чем покинуть рибосому. Точно так же тРНК на A / A-сайте принимает временную конформацию A / P-связывания перед оседанием на P / P-сайте, готовую к следующей аминокислоте для продолжения трансляции.

викторина
1. Какая из этих структур обнаружена на тРНК?A. Антикондон петляB. кодоныC. AATSD. Все вышеперечисленное
Ответ на вопрос № 1
верно. Только антикодонная петля обнаружена в тРНК. Кодоны обнаруживаются на мРНК, а AATS являются ферментами, а не частями молекулы РНК.
2. Какие из этих модифицированных остатков обнаружены в тРНК?A. псевдоуридинаB. тимидинаC. цитидинD. Все вышеперечисленное
Ответ на вопрос № 2
D верно. Трансферные РНК содержат ряд модифицированных оснований, и эти модификации необходимы для его правильного функционирования. Пока что они являются единственным классом молекул РНК, которые, как известно, содержат тимидин, обычно наблюдаемый только в ДНК. Псевдоуридин и цитидин обычно видны в T-плече тРНК.
3. Какое из этих модифицированных оснований происходит от аденина?A. аденозинB. ИнозинC. цитидинD. Все вышеперечисленное
Ответ на вопрос № 3
В верно. Инозин получен из аденина. Хотя технически можно сказать, что аденозин происходит из аденина, он не считается «модифицированным» основанием. Цитидиновый нуклеотид образуется, когда цитозин присоединяется к фосфорилированному рибозному сахару.
В результате чего образуется трнк
ГяРНК свойственна исключительно эукариотам, её нет у прокариот («доядерные» организмы, например бактерии и вирусы). Некоторые вирусы для хранения и передачи генетической информации следующим поколениям используют РНК вместо ДНК.
ГяРНК и её производное — информационная (или матричная) РНК переносят генетическую информацию от ядерной ДНК к цитоплазме.
Количество видов гяРНК равно количеству генов, так как она служит прямой копией кодирующих последовательностей генома. В процессе транскрипции РНК с ДНК ключевую роль играет фермент РНК-полимераза II. Информационная РНК образуется в результате процессинга гяРНК, при котором происходят вырезание некодирующих участков (интронов) и склеивание кодирующих экзонов. Таким образом, в состав иРНК входят кодирующая информация соответствующих видов гяРНК, а также фланкирующий лидерный и трейлерный участки, по этой причине она значительно короче.
Транспортная РНК
Каждая молекула тРНК состоит примерно из 75 связанных между собой нуклеотидов, образующих длинную цепь. В результате взаимодействия входящих в её состав оснований тРНК имеет конформационную структуру «клеверный лист», который затем скручивается в L-форму. Очень часто в состав тРНК помимо Ц, Г, А и У входит ряд редких оснований, некоторые из которых модифицированы путём метилирования. Важная особенность тРНК: «заряженная» молекула несёт на своём 3′-конце аминокислоту, а посередине конформационной структуры «клеверный лист» находятся три специфических основания, именуемые антикодоном. Последовательность оснований в антикодоне напрямую зависит от вида аминокислоты, прикреплённой к 3′-концу.
Так, например, тРНК, антикодон которой имеет последовательность 5′-ЦЦА-3′, может нести только аминокислоту триптофан. Следует отметить, что данная зависимость лежит в основе передачи генетической информации, носителем которой выступает тРНК.
Транскрипция молекул тРНК происходит с кодирующих её последовательностей в ДНК при участии фермента РНК-полимеразы III. Различают более 40 семейств тРНК, которые, в свою очередь, подразделяют на несколько видов.
Рибосомальная РНК
Существует несколько субъединиц рРНК, которые различаются по коэффициенту седиментации (осаждения), измеряемому в единицах Сведберга (S). Данный коэффициент зависит от скорости осаждения субъединиц при центрифугировании в насыщенной водной среде.
Каждая рибосома состоит из большой и малой субъединиц. Они содержат большое количество белков, синтезированных посредством трансляции иРНК, а также РНК, которая не подвергается трансляции. Термин «рибосомальная РНК» относят именно к нетранслируемому материалу. В малой субъединице находится 18S рРНК, а в большой — 4S, 5,8S и 28S рРНК.
Траскрипция рРНК с ДНК происходит при помощи двух дополнительных РНК-полимераз. РНК-полимераза I транскрибирует 5S, 5,8S и 28S в виде одного длинного 45S-тpaнскрипта, который затем разделяется на необходимые части. Таким образом обеспечивается равное количество молекул. В организме человека в каждом гаплоидном геноме присутствует примерно 250 копий последовательности ДНК, кодирующей 45S-транскрипт. Они расположены в пяти кластерных тандемных повторах в коротких плечах хромосом 13, 14, 15, 21 и 22.
Данные участки известны как ядрышковые организаторы, так как их транскрипция и последующий процессинг 45S-транскрипта происходят внутри ядрышка.
Не менее чем в трёх кластерах хромосомы 1 существует 2000 копий 5S-pPHK гена. Их транскрипция протекает в присутствии РНК-полимеразы III снаружи ядрышка. Затем они доставляются к местам сборки рибосом при помощи рибосомальных белков.
В рРНК насчитывают около 95 псевдоуридиновых участков, образованных посредством изомеризации уридина малой ядрышковой РНК.
Малая ядрышковая РНК. Малая (низкомолекулярная) ядрышковая РНК в основном участвует в направлении или проведении модификаций оснований в рРНК и малой ядерной РНК, таких, как, например, метилирование и псевдоуридинизация. Большинство малых ядрышковых РНК находятся в интронах других генов.
Сигналраспознающая РНК. Сигналраспознающая РНК распознаёт сигнальную последовательность белков, предназначенных для экспрессии, и участвует в их переносе через цитоплазматическую мембрану.
Микро-РНК. Существует примерно 200 микро-РНК человека длиной в 22 основания, производных расщепления рибонуклеазой Н их предшественников (двухцепочечных «зашпиленных» РНК) в соответствии с инвертированными повторами. Они контролируют трансляцию структурных генов путём комплементарного связывания с З’-концами нетранслируемых участков иРНК.
Митохондриальная РНК
Митохондриальная ДНК представляет собой непрерывную петлю и кодирует 13 полипептидов, 22 тРНК и 2 рРНК (16S и 23S). Большинство генов находятся на одной (тяжёлой) цепи, однако некоторое их количество расположено и на комплементарной ей лёгкой. При этом обе цепи транскрибируются в виде непрерывных транскриптов при помощи митохондриоспецифической РНК-полимеразы. Данный фермент кодируется ядерным геном. Длинные молекулы РНК затем расщепляются на 37 отдельных видов, а мРНК, рРНК и тРНК совместно транслируют 13 мРНК. Большое количество дополнительных белков, которые поступают в митохондрию из цитоплазмы, транслируются с ядерных генов.
У пациентов с системной красной волчанкой обнаруживают антитела к снурп-белкам собственного организма. Кроме того, считают, что определённый набор генов малой ядерной РНК хромосомы 15q играет важную роль в патогенезе синдрома Прадера—Вилли (наследственное сочетание олигофрении, низкого роста, ожирения, гипотонии мышц).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Транскрипция и трансляция

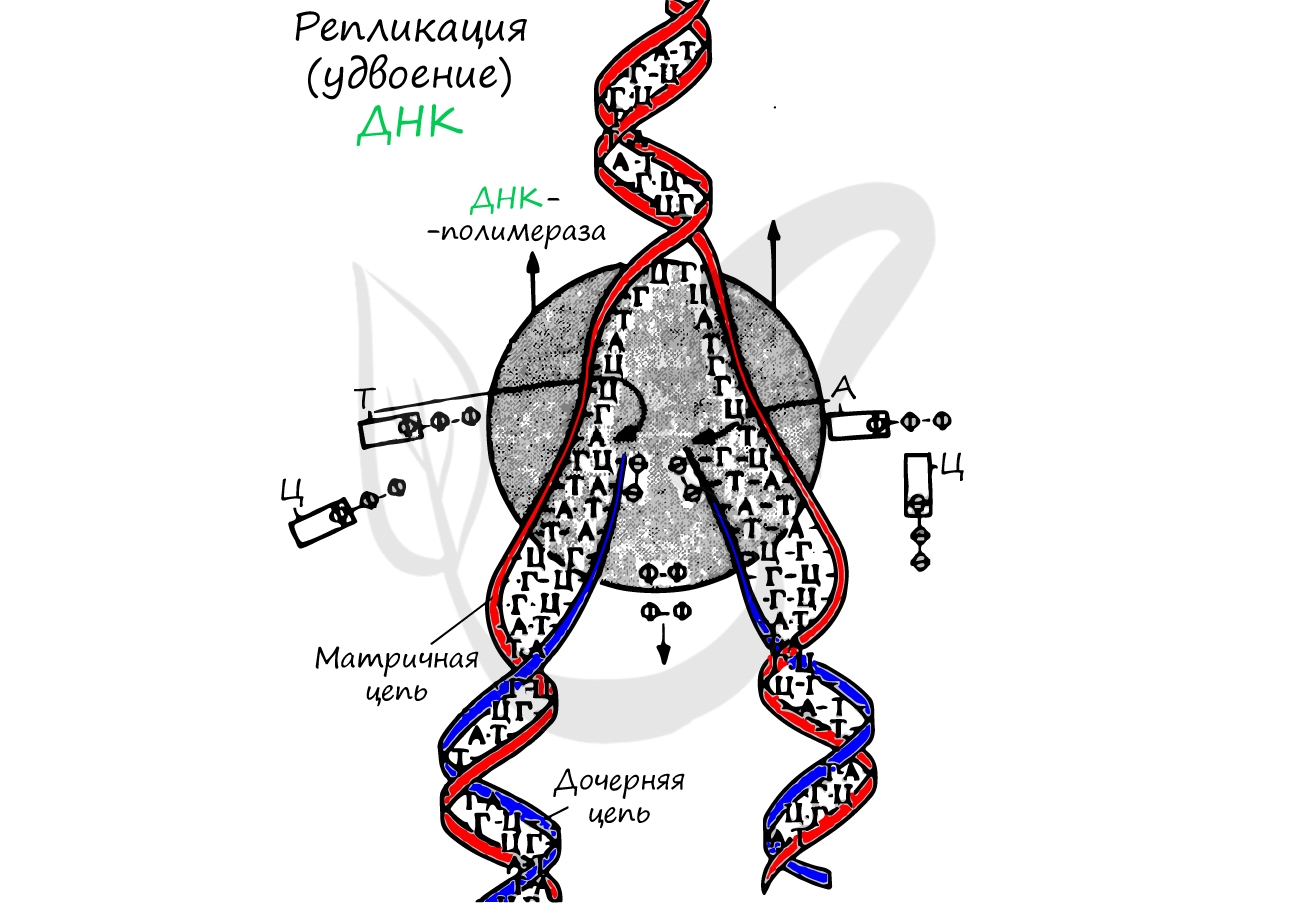
Удвоение ДНК происходит в синтетическом периоде интерфазы. При этом общее число хромосом не меняется, однако каждая из них содержит к началу деления две молекулы ДНК: это необходимо для равномерного распределения генетического материала между дочерними клетками.
Транскрпиция (лат. transcriptio — переписывание)
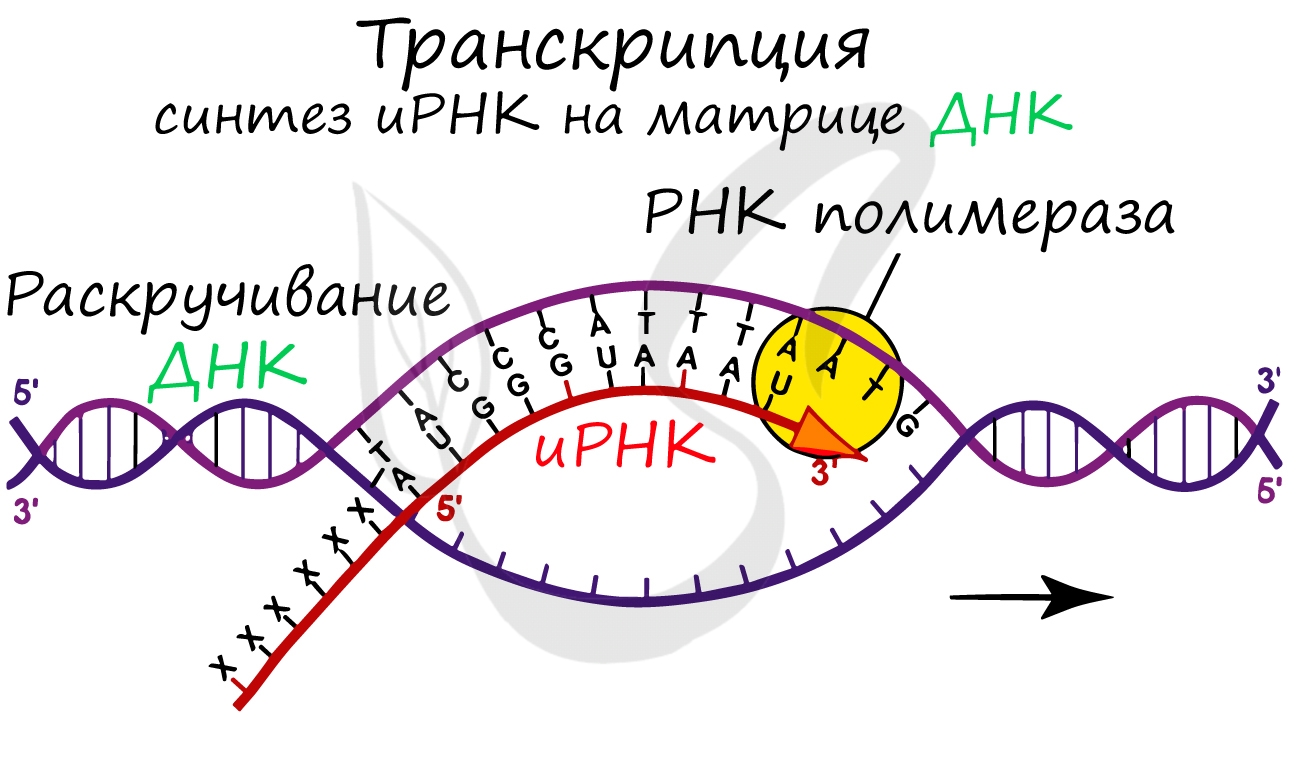
Образуется несколько начальных кодонов иРНК.
Нити ДНК последовательно расплетаются, освобождая место для передвигающейся РНК-полимеразы. Молекула иРНК быстро растет.
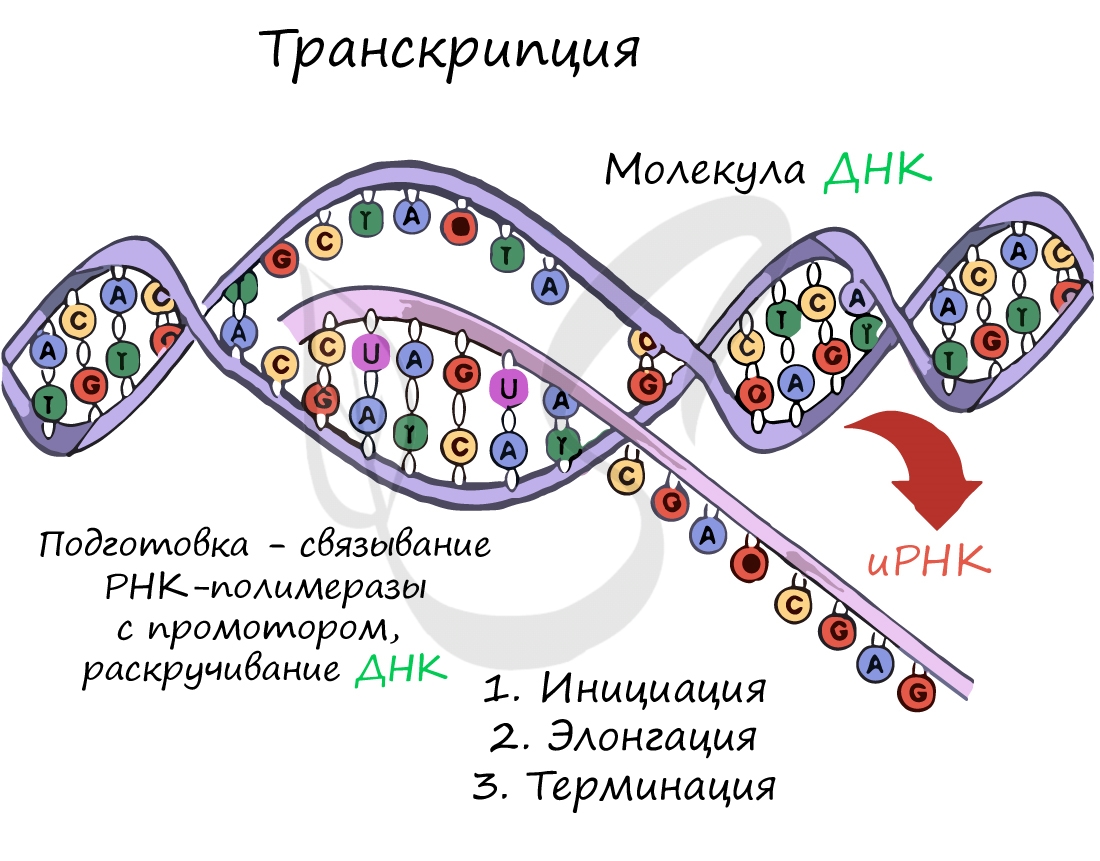
Трансляция (от лат. translatio — перенос, перемещение)
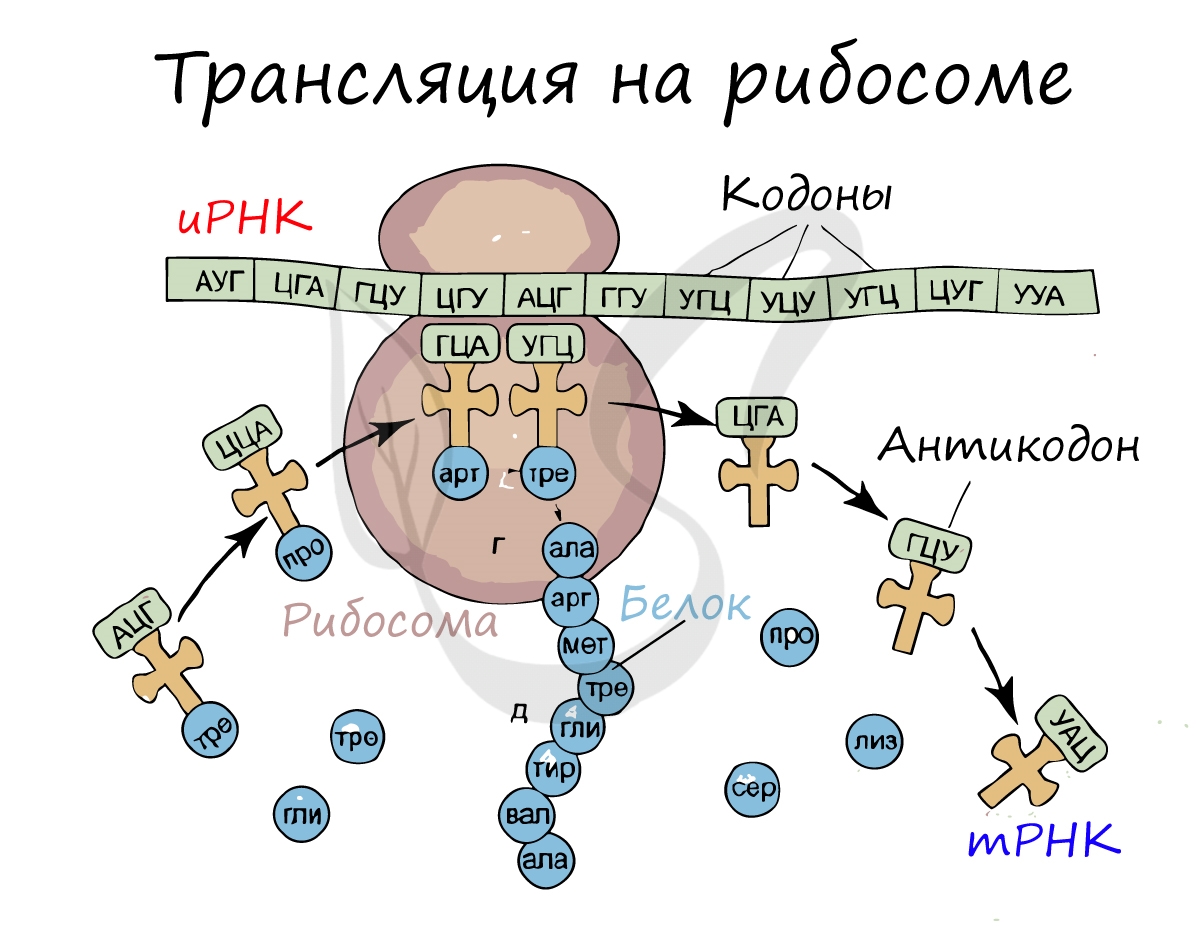
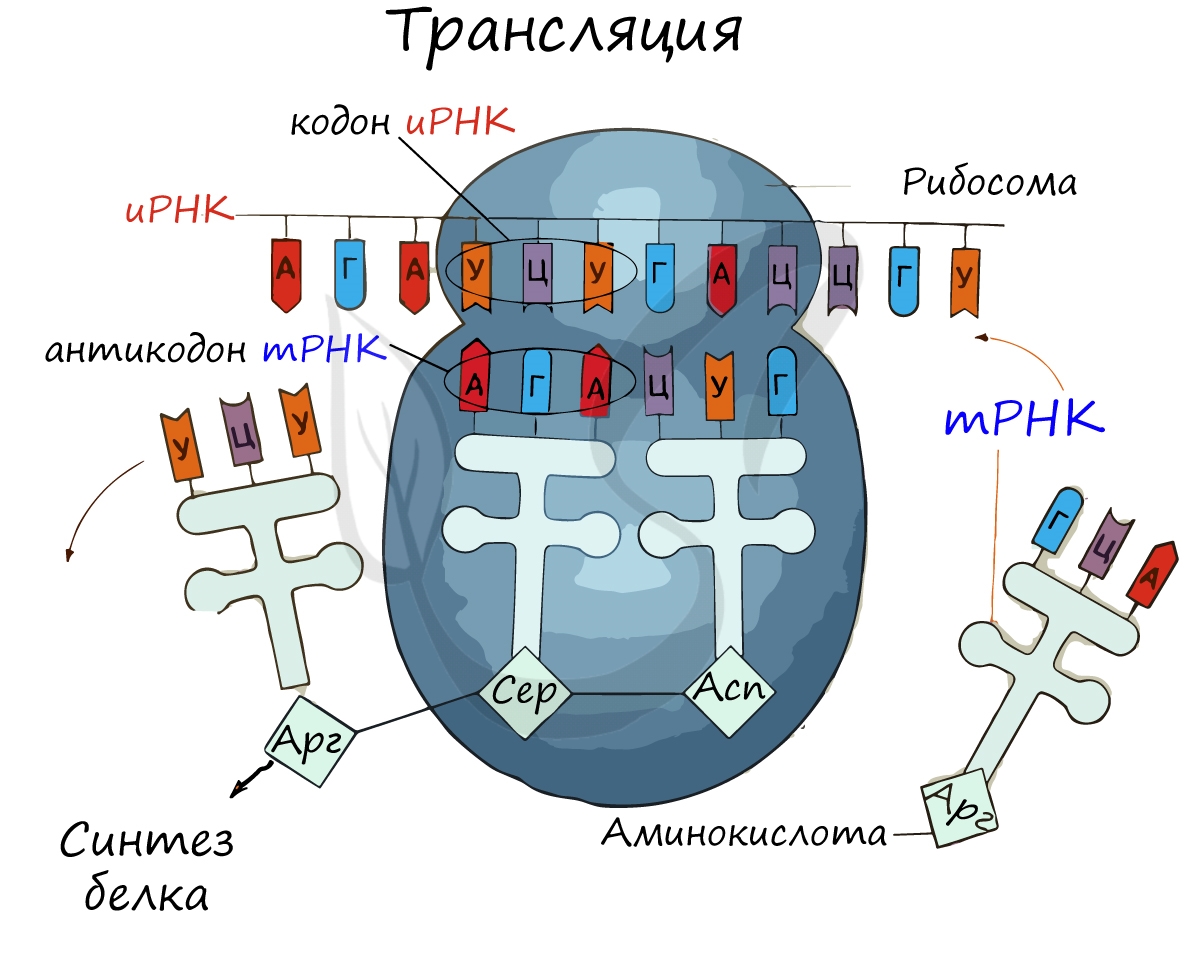
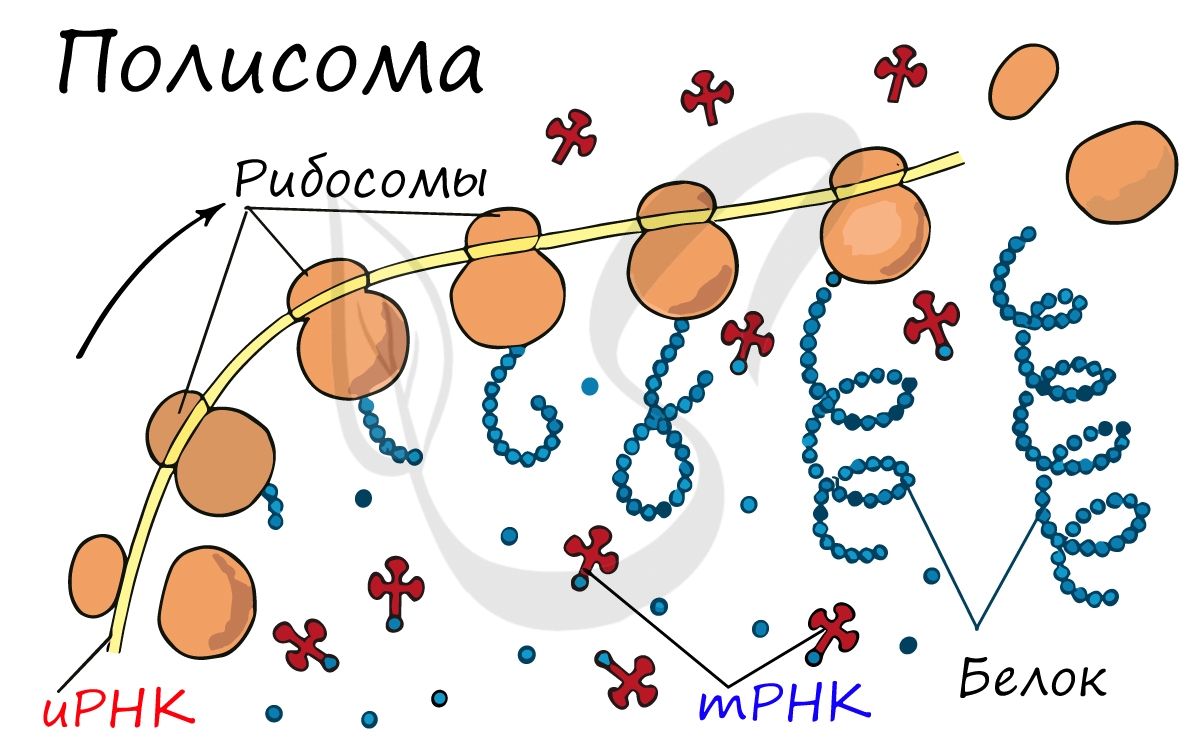
Рибосома делает шаг, и иРНК продвигается на один кодон: такое в фазу элонгации происходит десятки тысяч раз. Молекулы тРНК приносят новые аминокислоты, соответствующие кодонам иРНК. Аминокислоты соединяются друг с другом: между ними образуются пептидные связи, молекула белка растет.
Примеры решения задачи №1
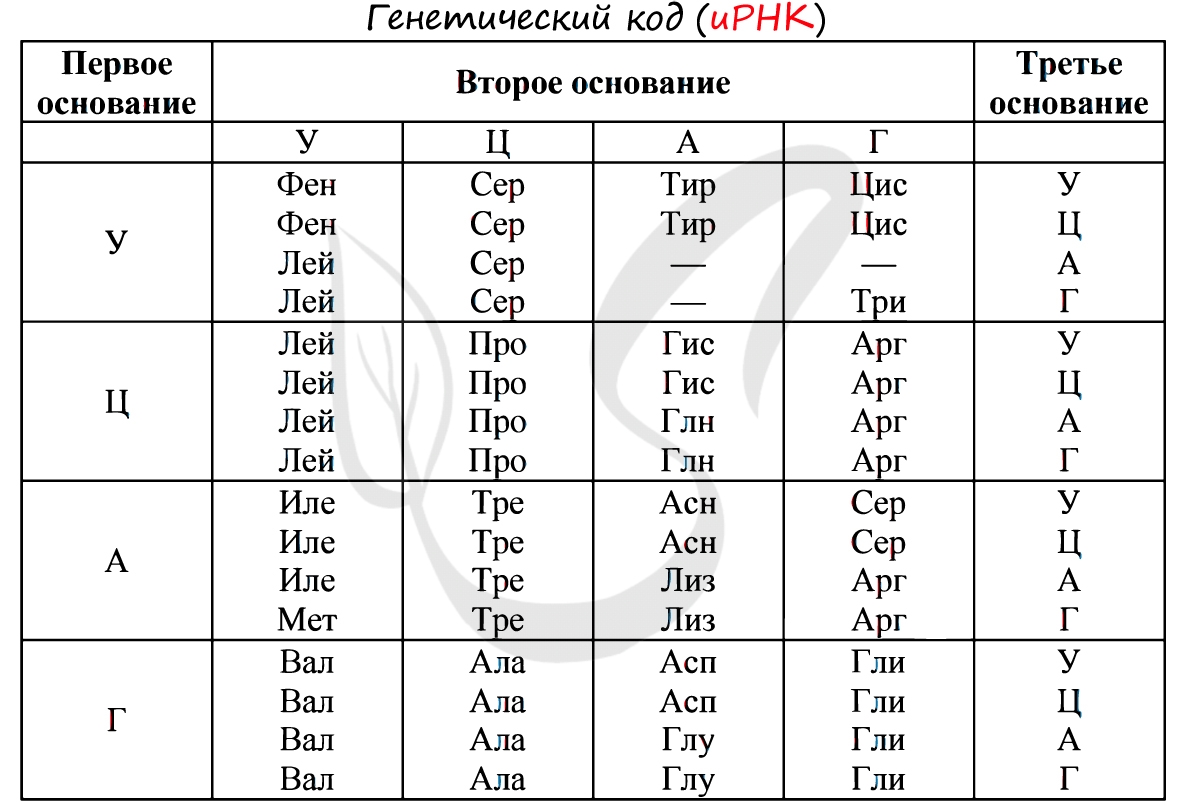
Без практики теория мертва, так что скорее решим задачи! В первых двух задачах будем пользоваться таблицей генетического кода (по иРНК), приведенной вверху.
«Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ЦГА-ТГГ-ТЦЦ-ГАЦ. Определите последовательность нуклеотидов во второй цепочке ДНК, последовательность нуклеотидов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка, используя таблицу генетического кода»

По принципу комплементарности мы нашли вторую цепочку ДНК: ГЦТ-АЦЦ-АГГ-ЦТГ. Мы использовали следующие правила при нахождении второй нити ДНК: А-Т, Т-А, Г-Ц, Ц-Г.
Вернемся к первой цепочке, и именно от нее пойдем к иРНК: ГЦУ-АЦЦ-АГГ-ЦУГ. Мы использовали следующие правила при переводе ДНК в иРНК: А-У, Т-А, Г-Ц, Ц-Г.
Зная последовательность нуклеотидов иРНК, легко найдем тРНК: ЦГА, УГГ, УЦЦ, ГАЦ. Мы использовали следующие правила перевода иРНК в тРНК: А-У, У-А, Г-Ц, Ц-Г. Обратите внимание, что антикодоны тРНК мы разделяем запятыми, в отличие кодонов иРНК. Это связано с тем, что тРНК представляют собой отдельные молекулы (в виде клеверного листа), а не линейную структуру (как ДНК, иРНК).
Пример решения задачи №2
«Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК, имеет следующую последовательность нуклеотидов: ТАГ-ЦАА-АЦГ-ГЦТ-АЦЦ. Установите нуклеотидную последовательность участка тРНК, который синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта тРНК в процессе биосинтеза белка, если третий триплет соответствует антикодону тРНК»

Пример решения задачи №3
Длина фрагмента молекулы ДНК составляет 150 нуклеотидов. Найдите число триплетов ДНК, кодонов иРНК, антикодонов тРНК и аминокислот, соответствующих данному фрагменту. Известно, что аденин составляет 20% в данном фрагменте (двухцепочечной молекуле ДНК), найдите содержание в процентах остальных нуклеотидов.

Теперь мы украсили теорию практикой. Что может быть лучше при изучении новой темы? 🙂
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Жизнь начиналась с РНК
Исследования нуклеиновых кислот являются одной из самых «горячих точек» в биологии. Благодаря уникальным свойствам РНК находят все более широкое применение в медицине и технике. Но знает об этом пока лишь узкий круг специалистов.
Рибонуклеиновой кислоте, иначе – РНК – не повезло. Она не пользуется такой широкой известностью, как ее близкий «родственник» – ДНК, несмотря на большое химическое сходство. Однако открытия последних двадцати лет радикально поменяли наши взгляды на роль и функции этих, как выяснилось, очень «умелых» молекул. Плодом этих открытий стала принципиально новая идея о том, что современной жизни предшествовал совершенно самодостаточный древний «мир РНК».
Как это обычно бывает, новое знание, расширяя горизонт, породило и массу новых вопросов. Каковы были механизмы «эволюции» в мире РНК? Зачем, откуда и как появились ДНК и белки? Как произошел переход от «мира РНК» к современному миру? О поисках, которые ведутся в этом направлении, читателям рассказывают академик Валентин Викторович Власов и его сын, кандидат химических наук, Александр Власов.
Почему в цикле статей, посвященных проблеме возникновения жизни, появляется статья об РНК, а не о других, более известных органических молекулах — ДНК или белках? Возможно, наши читатели слышали и об РНК, но вот что? Уверены, ничего примечательного — по одной простой причине: пока лишь специалисты-биологи знают, что именно РНК являются «волшебными» молекулами, давшими начало жизни. Что когда-то в древности, на только что остывшей Земле, возник и существовал загадочный «мир РНК»…
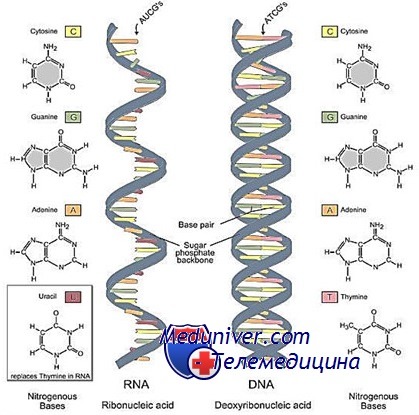
Прежде чем отправиться к «началу начал», давайте запасемся необходимыми знаниями о строении нуклеи-новых кислот — ДНК (дезоксирибонуклеиновой) и РНК (рибонуклеиновой). По своему химическому составу РНК является двойняшкой, хотя и не полным близнецом, ДНК, основного хранителя генетической информации в живой клетке. Нуклеиновые кислоты представляют собой полимерные макромолекулы, состоящие из отдельных звеньев — нуклеотидов. Скелетом макромолекулы являются молекулы пятиуглеродного сахара, соединенные остатками фосфорной кислоты. К каждой молекуле сахара присоединяется одно азотистое основание. Нуклеотиды, которые различаются между собою только разными азотистыми основаниями, обозначаются буквами A, U, G, C (в РНК) и A, T, G, C (в ДНК).
Честно говоря, насчет РНК никто не задумывался долгие годы. Существовала догма, что вот есть клетка, есть хромосомы, в которых есть ДНК — хранитель генетической информации.
В конце концов, на рибосомах синтезируются белки. А РНК — она где-то в промежутке, переносчик информации от ДНК — и только. А потом посыпались открытия, которые заставили совершенно по-другому взглянуть на РНК Главное отличие нуклеиновых кислот заключается в их углеводной компоненте. В РНК сахар — рибоза, а в ДНК — дезоксирибоза: там, где у ДНК имеется атом водорода (Н), у РНК стоит оксигруппа (ОН). Результаты таких незначительных, на неискушенный взгляд, различий поражают. Так, ДНК существуют в основном в форме всем известных жестких спиралей, в которых две цепи ДНК удерживаются вместе за счет образования водородных связей между комплементарными нуклеотидами.
РНК также могут формировать спирали из двух цепочек, похожие на спирали ДНК, однако в большинстве случаев РНК существуют в виде сложных структур-клубков. Структуры эти формируются не только за счет образования упомянутых водородных связей между разными участками РНК, но и благодаря оксигруппе рибозы, которая может образовывать дополнительные водородные связи и взаимодействовать с фосфорной кислотой и ионами металлов. Глобулярные структуры РНК не только внешне напоминают белковые структуры, но и приближаются к ним по свойствам: они могут взаимодействовать с самыми разными молекулами, как маленькими, так и полимерными.
Кого Считать «Живым»?
Почему же именно РНК мы называем праматерью ныне существующей жизни? Чтобы ответить на этот вопрос, давайте разберемся, где проходит граница между живым и неживым.
Поскольку над проблемой происхождения жизни работают ученые из разных областей, каждый оперирует терминами близкой ему науки. Химики обязательно вспомнят слово «катализатор», математики — «информация». Биологи будут считать живой систему, содержащую вещество (генетическую программу), которое может копироваться (или, по-простому, размножаться). При этом необходимо, чтобы в ходе такого копирования могли происходить некоторые изменения наследственной информации и возникать новые варианты систем, т. е. должна существовать возможность эволюции. Еще биологи обязательно заметят, что такие системы должны быть пространственно обособлены. Иначе возникшие более прогрессивные системы не смогут воспользоваться своими преимуществами, поскольку их более эффективные катализаторы и другие продукты будут беспрепятственно «уплывать» в окружающую среду.
Каким же образом первые молекулярные системы были обособлены от окружающей среды? Колонии молекул могли, например, удерживаться вместе за счет адсорбции на какой-нибудь минеральной поверхности или пылевых частицах. Однако возможно, что уже самые примитивные системы располагали, подобно современным живым клеткам, настоящей мембранной оболочкой. Дело в том, что такая «протоклетка» с липидной мембраной может образоваться очень просто. Многие молекулы с заряженными группами (например, жирные кислоты) в водной среде образуют микроскопические пузырьки — липосомы. Это слово должно быть хорошо известно прекрасной половине наших читателей: липосомы широко используются в косметических кремах — крохотные жировые капсулы начиняются витаминами и другими биологически активными веществами. А вот чем были наполнены древние «протоклетки»? Оказалось, что на роль «начинки» претендуют именно РНК.
РНК умеет все?
Жизнь, без сомнения, должна была начаться с образования «умелых» молекул, которые могли бы сами себя размножать и выполнять все другие «хозяйственные работы», необходимые для существования клетки. Однако на роль таких умельцев не подходит ни ДНК, ни белок. ДНК — отличный хранитель генетической информации, но сама себя размножать не умеет. Белки — непревзойденные катализаторы, но не могут работать в качестве «генетических программ». Возникает парадокс курицы и яйца: ДНК не может образоваться без белка, а белок — без ДНК. И только РНК, как выяснилось, может ВСЕ. Но не будем забегать вперед.
Рассмотрим давно известные функции РНК, связанные с работой (экспрессией) гена в клетке. При включении гена сначала происходит локальное расплетение ДНК и синтезируется РНК-копия генетической программы. В результате сложных обработок ее специальными белками получается матричная РНК (мРНК), которая и явля-ется программой для синтеза белка. Эта РНК переносится из ядра в цитоплазму клетки, где она связывается со специальными клеточными структурами — рибосомами, настоящими молекулярными «машинами» для синтеза белка. Белок синтезируется из активированных аминокислот, присоединенных к особым транспортным РНК (тРНК), причем каждая из аминокислот присоединена к своей специфической тРНК. Благодаря тРНК аминокислота фиксируется в каталитическом центре рибосомы, где она «пришивается» к синтезируемой белковой цепи. Из рассмотренной последовательности событий видно, что молекулы РНК играют ключевую роль в декодировании генетической информации и биосинтезе белка.
Чем больше углублялись в изучение различных биосинтетических процессов, тем чаще обнаруживали ранее неизвестные функции РНК. Оказалось, что кроме процесса транскрипции (синтеза РНК путем копирования участка ДНК) в ряде случаев, наоборот, может происходить синтез ДНК на РНК-матрицах. Этот процесс, названный обратной транскрипцией, используют в ходе своего развития многие вирусы, в том числе печально известные онкогенные вирусы и ВИЧ-1, вызывающий СПИД.
Таким образом, выяснилось, что поток генетической информации не является, как первоначально считалось, однонаправленным — от ДНК к РНК. Роль ДНК как изначально главного носителя генетической информации стала подвергаться сомнению. Тем более что многие вирусы (гриппа, клещевого энцефалита и другие) вообще не используют ДНК в качестве генетического материала, их геном построен исключительно из РНК. А далее посыпались одно за другим открытия, которые заставили совершенно по-другому взглянуть на РНК.
На Все «Молекулы» Мастер
Наиболее удивительным было открытие каталитической способности РНК. Прежде считалось, что катализировать реакции умеют только белки, ферменты. Ученые, например, никак не могли выделить ферменты, осуществляющие разрезание и сшивание некоторых РНК. После длительных исследований выяснилось, что РНК прекрасно справляются с этим сами. Структуры РНК, действующие подобно ферментам, назвали рибозимами (по аналогии с энзимами, белками-катализаторами). Вскоре было обнаружено множество разнообразных рибозимов. Особенно широко их используют для манипулирования своими РНК вирусы и другие простые инфекционные агенты. Таким образом, РНК оказались мастерами на все руки: они могут выступать в роли носителей наследственной информации, могут служить катализаторами, транспортными средствами для аминокислот, образовывать высокоспецифичные комплексы с белками.
Окончательная уверенность в том, что «мир РНК» действительно существовал, наступила после выявления деталей строения кристаллов рибосом методом рентгеноструктурного анализа. Ученые рассчитывали обнаружить там белок, катализирующий сшивание аминокислот в белковую последовательность. Каково же было их удивление, когда выяснилось, что в каталитическом центре рибосом белковых структур нет совсем, что он полностью построен из РНК! Оказалось, что все ключевые стадии биосинтеза белка осуществляются молекулами РНК. Точка в дискуссии о возможности существования «мира РНК» как особой стадии биологической эволюции была поставлена.
Конечно, полную картину еще предстоит реконструировать — осталось много нерешенных вопросов. Например, в современной клетке активацию аминокислот и их присоединение к соответствующим тРНК осуществляют специфичные белки-ферменты. Возникают вопросы: могла ли эта реакция осуществляться без участия белков, только с помощью РНК? Могли ли сами РНК катализировать синтез РНК из нуклеотидов или присоединение азотистых оснований к сахару? В общем-то, после открытия рибозимов такие потенциальные способности РНК уже не вызывали особых сомнений. Но наука требует, чтобы гипотезы экспериментально подтверждались.
Дарвиновская Эволюция в Пробирке
Хороший метод зачастую позволяет осуществить революцию в науке. Именно так можно сказать о методе полимеразной цепной реакции (ПЦР), который позволяет размножать нуклеиновые кислоты в неограниченных количествах. Кратко опишем суть метода. Для размножения ДНК в методе ПЦР используются ферменты ДНК- полимеразы, т. е. те самые ферменты, которые при размножении клеток синтезируют из активированных мономеров-нуклеотидов комплементарные цепочки ДНК.
При методе ПЦР в пробирку с ДНК вносят смесь активированных нуклеотидов, фермент ДНК-полимеразу и так называемые праймеры — олигонуклеотиды, комплементарные концам размножаемой ДНК. При нагревании раствора цепи ДНК расходятся. Затем, при охлаждении, с ними связываются праймеры, образуя короткие фрагменты спиральных структур. Фермент присоединяет к праймерам нуклеотиды и собирает цепочку, комплементарную цепочке исходной ДНК. В результате реакции из одной двуцепочечной ДНК получается две. Если повторить процесс, получится четыре цепочки, а после n повторений — 2 n молекул ДНК. Все очень просто.
Изобретение ПЦР и разработка методов химического синтеза ДНК позволили создать потрясающую технологию молекулярной селекции. Принцип молекулярной селекции тоже прост: сначала синтезируется множество молекул, обладающих разными свойствами (так называемая молекулярная библиотека), а затем из этой смеси отбираются молекулы с желаемым свойством.
Библиотеки нуклеиновых кислот — это смеси молекул, имеющих одинаковую длину, но отличающихся последовательностью нуклеотидов. Получить их можно в том случае, если при химическом синтезе на авто-матическом синтезаторе добавлять на каждой стадии удлинения нуклеотидной последовательности одно-временно все четыре нуклеотида. Каждый из них будет включаться в растущую нуклеиновую кислоту с равной вероятностью, в результате чего на каждом этапе присоединения будет получаться 4 варианта последовательностей. Если таким образом синтезировать нуклеиновую кислоту длиной в n звеньев, то разнообразие полученных молекул составит 4 в степени n. Поскольку обычно используются участки длиной 30—60 мономеров, то в результате синтеза получается от 4 30 до 4 60 разных молекул! Цифры, привычные разве что для астрономов.
Так как в зависимости от состава нуклеиновые кислоты сворачиваются в разные пространственные структуры, синтез статистических последовательностей дает огромное множество молекул, различающихся по свойствам. С образовавшихся ДНК — с помощью фермента РНК-полимеразы — считывается РНК. В результате получается библиотека уже одноцепочечных РНК. Далее производится процедура отбора: раствор РНК пропускается через колонку, в которой находится нерастворимый носитель с химически присоединенными молекулами-мишенями, чтобы «выловить» так называемый будущий аптамер, т. е. РНК, способную связывать определенные молекулы. Затем колонку промывают для удаления несвязавшихся РНК, а затем смывают РНК, задержавшиеся на колонке за счет связывания с целевыми молекулами (это можно сделать, например, нагревая колонку).
С выделенных РНК с помощью обратной транскрипции делают ДНК-копии и получают из них обычные двуцепочечные молекулы ДНК. С последних же можно считывать искомые РНК-аптамеры, а затем — размножать их методом ПЦР в неограниченных количествах. Конечно, так происходит в идеальном случае, на практике все получается сложнее. Обычно исходный препарат РНК содержит огромный избыток «по-сторонних» молекул, избавиться от которого трудно. Поэтому полученную РНК вновь и вновь пропускают через колонку, чтобы выделить РНК, образующие самые прочные комплексы с целевыми молекулами.
С помощью такого метода были получены тысячи разных РНК-аптамеров, которые образуют специфические комплексы с различными органическими соединениями и молекулами.
Рассмотренная схема молекулярной селекции может быть применена для получения молекул с любыми свойствами. Например, были получены РНК, способные катализировать реакции синтеза РНК и белков: присоединение азотистых оснований к рибозе, полимеризацию активированных нуклеотидов на цепочках РНК, присоединение аминокислот к РНК. Эти исследования еще раз подтвердили, что в условиях предбиологической эволюции из случайных полимеров могли возникать молекулы РНК
со специфическими структурами и функциями.
Делайте Ваш Заказ!
Метод молекулярной селекции обладает очень большими возможностями. С его помощью можно решать задачи поиска нужных молекул даже в том случае, если исходно нет идеи, как такие молекулы должны быть устроены. Однако, если придумать процедуру отбора, их можно выделить по принципу требуемых свойств, а затем уже заняться и вопросом, как эти свойства достигаются. Продемонстрируем это на примере выделения РНК, способных связываться с клеточными мембранами и модулировать их проницаемость.
Древние рибоциты должны были поглощать «питательные» вещества из окружающей среды, удалять продукты метаболизма и делиться в ходе размножения.
И все эти процессы требуют управления проницаемостью мембран. Поскольку мы полагаем, что никаких других функциональных молекул, кроме РНК, в рибоцитах не было, какие-то РНК обязательно должны были взаимодействовать с мембранами. Однако с химической точки зрения они совершенно не подходят для роли регуляторов проницаемости мембран.
Мембраны современных клеток и липосом, построенные из жирных кислот, несут отрицательный заряд. Поскольку РНК также заряжены отрицательно, то по закону Кулона они должны отталкиваться от липидной поверхности и тем более не могут проникать в глубь липидного слоя. Единственный известный способ взаимодействия нуклеиновых кислот с поверхностью мембран — через двухзарядные ионы металлов. Эти положительно заряженные ионы могут играть роль мостиков, располагаясь между отрицательно заряженными группами на поверхности мембраны и фосфатными группами нуклеиновой кислоты. Поскольку такие мостиковые взаимодействия достаточно слабые, с мембраной может связаться только очень большая нуклеиновая кислота благодаря множеству слабых связей с поверхностью мембраны. Так маленькие враги привязали Гулливера к земле множеством тоненьких веревок.
Тут и помог исследователям метод молекулярной селекции. Из библиотеки РНК удалось выделить не-сколько молекул, которые очень успешно связывались с мембранами, а при достаточно высокой концентрации — даже разрывали их! Эти РНК обладали необычными свойствами. Они как бы помогали друг другу: смесь молекул разных сортов связывалась с мембранами гораздо лучше, чем молекулы одного сорта. Все стало ясным после изучения вторичных структур этих РНК. Оказалось, что в них имеются петли с комплементарными участками. За счет этих участков «мембранные» РНК могут формировать комплексы-сообщества, которые способны образовывать множественные контакты с мембраной и делать то, что одной молекуле РНК не под силу.
Этот селекционный эксперимент подсказал, что у РНК есть дополнительный способ приобретения новых свойств путем образования сложных надмолекулярных комплексов. Этот механизм мог использоваться и для удерживания эволюционирующих систем РНК в виде колоний на поверхностях еще до того, как эти системы обзавелись изолирующей мембраной.
«Мир РНК»: Был, Есть и Будет!
Множество данных свидетельствует о том, что «мир РНК» действительно существовал. Правда, не совсем ясно — где. Некоторые специалисты полагают, что начальные этапы эволюции происходили не на Земле, что на Землю были занесены уже функционально активные системы, которые приспособились к местным условиям. Однако с химической
и биологической точки зрения это не меняет сути дела. В любом случае остается загадкой — в результате каких процессов в окружающей среде рибоциты образовались и за счет каких компонентов существовали. Ведь требуемые для жизни рибоцитов нуклеотиды — сложные молекулы. Трудно представить, что эти вещества могли образовываться в условиях пребиотического синтеза.
Вполне возможно, что древние РНК значительно отличались от современных. К сожалению, следов этих древних РНК экспериментально обнаружить нельзя, речь идет о временах, удаленных от нас на миллиарды лет. Даже скалы тех времен давно «рассыпались в песок». Поэтому речь может идти только об экспериментальном моделировании процессов, которые могли протекать на самых ранних стадиях молекулярной эволюции.
Почему произошел переход от «мира РНК» к современному миру? Белки, располагающие гораздо большим набором химических групп, чем РНК, являются лучшими катализаторами и структурными элементами. По-видимому, некоторые древние РНК стали использовать белковые молекулы в качестве «орудий труда». Такие РНК, способные к тому же синтезировать для своих целей полезные молекулы из окружающей среды, получали преимущества в размножении. Естественным путем отбирались соответствующие аптамеры и рибозимы.
А затем эволюция сделала свое дело: возник аппарат трансляции, и постепенно ответственность за катализ перешла к белкам. Орудия оказались столь удобными, что вытеснили своих «хозяев» из многих сфер деятельности.
Читатель вправе спросить: а зачем вообще нужно исследовать эволюцию РНК, ведь древний «мир РНК» исчез? Неужели только ради «чистого искусства», удовлетворения интересов фанатичных исследователей? Однако, не зная прошлого, нельзя понять настоящее. Изучение эволюции и возможностей РНК может подсказать новые направления поиска процессов, протекающих в современных живых клетках. Например, совсем недавно были обнаружены мощные системы регуляции активности генов с участием двуцепочечных РНК, с помощью которых клетка защищает себя от вирусных инфекций. Эта древняя система клеточной защиты, вероятно, скоро найдет применение в терапии.
Поэтому неудивительно, что в наше время исследования нуклеиновых кислот продолжают оставаться одной из самых «горячих точек» в молекулярной биологии. Благодаря уникальным свойствам РНК находят все более широкое применение в медицине и технике. Возникший в незапамятные времена «мир РНК» будет не только продолжать незримо существовать
в наших клетках, но и возрождаться в виде новых биотехнологий.
Редакция благодарит сотрудников Института химической биологии и фундаментальной медицины
СО РАН к. х. н. В. В. Коваля, к. х. н. С. Д. Мызину и к. х. н. А. А. Бондаря за помощь в подготовке статьи



