Научная электронная библиотека

Юров И. Ю., Ворсанова С. Г., Воинова В. Ю., Чурносов М. И., Юров Ю. Б.,
3.4. Деление клетки
Все клетки человека проходят через цикл деления (клеточный цикл). Известны два типа деления клетки: митотческое и мейотическое, которые представлены на рисунках 6 и 7. Митоз – деление клетки, за счет которого достигается воспроизведение соматических клеток в организме. Мейоз – деление клеток, в ходе которого получаются клетки с редуцированным гаплоидным (n) набором хромосом (половые клетки). Среднее время клеточного цикла у млекопитающих составляет 17–18 часов. Клеточный цикл разделен на четыре основных стадии: G1, S (фаза синтеза ДНК), G2 и митоз (М). Первые три фазы представляют собой интерфазный период или интерфазу. Клетки, которые не делятся, находятся на стадии покоя G0.
Во время стадии G1 (примерно 9 часов) хромосомы имеют вид одиночных хроматид, клетка метаболически активна и в ней происходит синтез белков. Если клетка, находящаяся на G1 стадии, не подвергается последующему делению, то это состояние соответствует G0. Фаза синтеза (или S фаза) длится около 5 часов и характеризуется процессом репликации хромосомной ДНК. На этой стадии хромосомы состоят из двух идентичных сестринских хроматид. G2 стадия длится примерно 3 часа. Во время этой стадии клетка готовится к процессу деления. Завершение G2 соответствует концу интерфазы. Митоз длится не более 1–2 часов и является процессом образования двух генетически идентичных дочерних клеток. В свою очередь, митоз также делится на 4 стадии: профаза, метафаза, анафаза и телофаза (рис. 6).
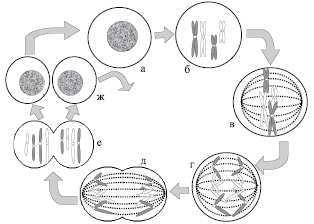
Рис. 6. Митоз. Схематическое изображение двух пар хромосом во время митоза: а – интерфаза; б – профаза; в – метафаза; г – анафаза; д – телофаза; е – цитокинез; ж – интерфазы разделившихся клеток
Профаза – стадия, во время которой происходит постепенная конденсация (уплотнение) и спирализация хромосом, в результате чего, они имеют вид дискретных структур. Во время профазы образуется веретено деления (двуполюсное веретено, состоящее из пучков микротрубочек, которые тянутся от одного полюса к другому).
Метафаза – стадия, характеризующаяся тем, что полностью осуществляется присоединение хромосом к нитям веретена, и хромосомы собираются в экваториальной плоскости клетки, находящейся на одинаковом расстоянии от обоих полюсов веретена. На этой стадии хромосомы достигают максимальной конденсации.
Анафаза – стадия разделения сестринских хроматид и их расхождения к противоположным полюсам веретена.
Телофаза – последняя стадия митоза. Она начинается, когда все сестринские хроматиды доходят до полюсов веретена. В ходе данной фазы митоза происходит восстановление клеточного ядра и внутриядерных структур. За ней, как правило, следует цитокинез – разделение двуядерной клетки на две с одним ядром в каждой (рис. 6).
Мейоз – деление клеток, в ходе которого получаются клетки с редуцированным гаплоидным набором хромосом (половые клетки). Данный процесс включает в себя две фазы клеточного деления: мейоз 1 и мейоз 2. В ходе мейоза происходит редукция диплоидного хромосомного набора (2n) до гаплоидного (n).
Мейоз 1, в свою очередь также делится на несколько стадий: профаза I, метафаза I, анафаза I и телофаза I (рис. 7). Профаза I является комплексной стадией, которая, в свою очередь, делится на несколько этапов:
лептотена: 46 хромосом состоят из 2-х хроматид и начинают коденсироваться; эта стадия характеризует начало мейоза;
зиготена: гомологичные хромосомы спариваются своими участками – этот этап деления называется синапсом; в результате образуется тройственная структура – синаптонемальный комплекс;
пахитена: стадия завершения синапса, спаренные гомологи, состоящие из 4-х хроматид, формируют бивалент; во время этой стадии происходит кроссинговер – обмен последовательностями ДНК хромосом между несестринскими хроматидами бивалентов. Результатом этого процесса является рекомбинация генетического материала между гомологичными хромосомами, создающая новые комбинации генов в дочерних клетках;
диплотена: на этой стадии хромосомы отталкиваются друг от друга до тех пор, пока гомологи не будут соединены только участками, подверженными кроссинговеру. Такие участки называются хиазмами;
диакинез: хромосомы претерпевают наибольшее сжатие во время этой последней стадии профазы I.
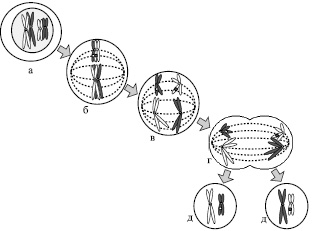
Рис. 7. Мейоз I. Схематическое изображение двух пар хромосом во время мейоза I: а – профаза I; б – метафаза I; в – анафаза I; г – телофаза I; д – клетки, образующиеся в результате первого мейоза
Метафаза I характеризуется исчезновением ядерной мембраны и образованием мейотического веретена деления. Биваленты выравниваются по экваториальной плоскости клетки и их центромеры случайным образом ориентируются к противоположным полюсам. Во время анафазы I биваленты разделяются и расходятся к противоположным полюсам. В ходе телофазы I каждая хромосома из двух гаплоидных наборов достигает противоположных полюсов, и образуются две дочерние клетки, в каждой из которых по 23 хромосомы, состоящие из 2-х хроматид.
Мейоз 2 практически идентичен митотическому делению за исключением того, что в данном случае делящиеся клетки имеют гаплоидный хромосомный набор. Хромосомы выравниваются по экваториальной плоскости клетки на стадии метафазы II, хроматиды разделяются и расходятся к противоположным полюсам на стадии анафазы II, цитокинез происходит на стадии телофазы II. В результате митотического деления (мейоза 1 и 2), как правило, образуются 4 дочерние клетки с гаплоидным набором хромосом, каждая из которых генетически отличается друг от друга за счет процесса кроссинговера и случайного расхождения гомологичных хромосом.
Научная электронная библиотека
Юров И. Ю., Ворсанова С. Г., Воинова В. Ю., Чурносов М. И., Юров Ю. Б.,
1.1. Хромосомы человека
Хромосомный набор человека, определяемый как кариотип – совокупность данных о структуре, размерах и количестве митотических хромосом, – установлен в начале 60-х годов прошлого века. Ещё в 1888 году Г. Вальдеер (H. Waldeyer, 1836–1921 гг.) ввёл термин «хромосома» для обозначения окрашенных нитевидных структур, видимых в ходе стадий деления клетки (митоза). Характерные особенности строения каждой хромосомы человека определяются, как известно, положением в ней центромеры – важнейшей структуры, которая в делении клетки (митозе) соединяется с нитями веретена и определяет расхождение сестринских хроматид к противоположным полюсам клетки. Метафазная хромосома состоит из двух хроматид (сестринские хроматиды) и центромеры, при помощи которой они соединяются. В районе центромеры хромосома сужена, две её хроматиды сближены, и этот район в теле хромосомы образует первичную перетяжку. Центромера делит хромосому на два плеча (короткое и длинное). По положению центромеры и первичной перетяжки среди хромосом человека различают метацентрические хромосомы, у которых центромера расположена в середине хромосомы (медианно) и делит её на два равных по длине плеча; субметацентрические хромосомы, в которых центромера расположена субмедианно и делит хромосому на два плеча неравной длины; и акроцентрические хромосомы, у которых центромера расположена почти на конце хромосомы (терминально), отделяя от длинного очень короткое плечо. У некоторых хромосом на коротком плече двух хроматид на красящейся тонкой нити располагаются маленькие хроматические тельца – спутники. Участки на концах хромосомы называются теломерами. Структуры в виде точек прикрепления нитей митотического веретена к центромерам называются кинетохорами. Плечи некоторых хромосом содержат перетяжки, называемые «вторичными» (например, хромосомы 1, 9, 16). Диплоидный набор человека, состоящий из 46 хромосом, составлен из 23 пар гомологичных хромосом – гомологов (отцовского и материнского происхождения): 22 пары аутосом и плюс половые хромосомы (гоносомы) – ХХ у женщин или ХY у мужчин. Гомологичные хромосомы, как правило, сходны между собой в размерах и строении, хотя могут встречаться некоторые отклонения от каждого показателя, и это носит название «гетероморфизм хромосом». Термин «кариотип» рекомендуется применять к систематизированному набору хромосом отдельной клетки человека. Существует также термин «идиограмма», который сохраняется для представления кариотипа в виде схемы, построенной на основании измерений хромосом большого числа клеток. Хромосомы пронумерованы серийно от 1 до 22 в соответствии с их длиной, а также с другими особенностями их строения, допускающими идентификацию. Половые хромосомы (гоносомы) не имеют номеров и обозначаются как Х и Y. Следует отметить, что термины и «кариотип», и «идиограмма», получившие международное признание и распространение, принадлежат русским цитологам: «идиограмма» – С.Г. Навашину (1857–1930 гг.) в 1921 году и «кариотип» – Г.А. Левитскому (1878–1942 гг.) в 1924 году. В фазах деления – метафазах и прометафазах (см ниже) – хромосомы можно увидеть в световом микроскопе как дискретные удлинённые структуры длиной от 2 до 11 мкм. На рисунках 1 и 2 представлены мужской и женский кариотипы человека.

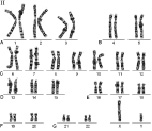
Рис. 1. Мужской кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
Как указано выше, кариотип человека состоит из 46 хромосом, которые нумеруются от 1 до 22 (аутосомы) и делятся на 7 групп, – A, B, C, D, E, F, G и половые хромосомы (гоносомы) X и Y (рис. 1 и 2).
К первой группе А относятся хромосомы 1, 2 и 3, которые хорошо отличаются друг от друга. Хромосома 1 (размер – 11 мкм) – метацентрическая, содержит вторичную перетяжку в околоцентромерном участке длинного плеча. Хромосома 2 (10,8 мкм) по размерам почти равна хромосоме 1 и является субметацентрической. Хромосома 3 (размер – 8,3 мкм) – практически метацентрическая.
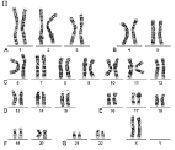
Рис. 2. Женский кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
К группе В относятся хромосомы 4 и 5 (размер – 7,7 мкм каждая) – это крупные субметацентрические хромосомы, которые не отличаются друг от друга при рутинном окрашивании ни размером, ни положением центромер.
К группе С относятся хромосомы с 6 по 12 и Х. В основном, это субметацентрические хромосомы крупных и средних размеров. Наиболее крупные хромосомы из группы С – 6, 7 и Х (6,8–7,2 мкм). Хромосома Х является половой хромосомой (гоносомой). Хромосома 7 более метацентрична, чем хромосома 6. Хромосомы 8 и 9 – практически одинаковы по размеру (5,8 мкм). Хромосома 8 метацентричнее хромосомы 9, которая характеризуется регулярной вторичной перетяжкой в прицентромерном районе длинного плеча.
К группе D относятся хромосомы 13, 14 и 15 (4,2 мкм) – средних размеров акроцентрические хромосомы с почти терминальным расположением центромеры. Эти хромосомы между собой не различаются ни по размерам, ни морфологически после рутинного окрашивания. Короткое плечо всех трёх пар хромосом может формировать спутники (рис. 1, 2 и 5).
К группе Е относятся хромосомы 16, 17 и 18. Хромосома 16 (размер – 3,6 мкм) – сравнительно небольшая метацентрическая хромосома, содержащая вторичную перетяжку в длинном плече. Хромосома 17 (размер – 3,5 мкм) – сравнительно короткая субметацентрическая хромосома. Хромосома 18 (размер – 3,2 мкм) – самая короткая субметацентрическая хромосома.
Группа F представлена хромосомами 19 и 20 (размеры – 2,9 мкм). Это короткие метацентрические хромосомы, которые не отличаются между собой без дифференциального окрашивания по длине.
Хромосомы 21 и 22 (2,8 мкм) относятся к группе G. Это наиболее короткие акроцентрические хромосомы в кариотипе, которые обладают способностью формировать спутники на коротком плече (рис. 1, 2 и 5).
Хромосома Y (2,3 мкм) является маленькой акроцентрической хромосомой, сравнимой по размерам с хромосомами 21 и 22, но не имеющая спутников.
Важнейшая работа по созданию общей системы обозначения и классификации хромосом человека, представляемая в виде отдельной книги, проводилась регулярно из года в год, куда в последние десятилетия включались и результаты использования современных молекулярно-цитогенетических методов. Первые результаты работы учёных по номенклатуре и классификации хромосом человека были представлены в 1960 году в Денвере, в Университете штата Колорадо (США). Работа комиссии была проведена при поддержке американского онкологического общества. В состав комиссии входили известнейшие учёные: 14 учёных-цитологов и 3 учёных-генетика. Результаты были опубликованы в Денвере в том же 1960 году в ряде журналов в виде документа (книги), названного как «Стандартная система номенклатуры митотических хромосом человека». Впоследствии был создан комитет по номенклатуре хромосом человека. Комиссия и комитет периодически собирались для работы и, по мере развития цитогенетических, а в последние десятилетия и молекулярно-цитогенетических исследований, вносили поправки и дополнения в существующую классификацию. Работа комиссий проводилась регулярно в различных странах и городах. Комитет последовательно собирался в Париже, Чикаго, Мехико, Лэйк-Плэсиде, Эдинбурге, Стокгольме, где было решено разработать и опубликовать унифицированный вариант номенклатуры хромосом человека, включающий основные положения первых совещаний (Денвер, Лондон, Чикаго, Париж). Результаты работы комитета публиковались в различных издательствах. Этот важный документ получил название «Международная система номенклатуры хромосом человека» – «An International System for Human Cytogenetic Nomenclature» (ISCN). В дальнейшем поправки и дополнения в документе делались, учитывая новые разработанные технологии, включая молекулярно-цитогенетические, и докладывались на различных конференциях. Последний документ – «ISCN 2016» – An International System for Human Cytogenetic Nomenclature» опубликован в 2016 году. Таким образом, в одном документе даётся полная номенклатура хромосом в норме и при хромосомных синдромах и аномалиях, исходя из цитогенетических и современных молекулярно-цитогенетических технологий (см рекомендуемую литературу).
Следует отметить, что хромосомные синдромы и аномалии связаны с хромосомными (геномными) мутациями (аномалиями) в виде различных структурных перестроек хромосом или с изменением их числа (n). Численные изменения хромосом могут быть двух типов: полиплоидии – умножение хромосомного набора (3n, 4n и т. д.) или генома, кратное гаплоидному числу хромосом; анеуплоидии – увеличение или уменьшение числа хромосом, некратное гаплоидному. Структурные хромосомные (геномные) перестройки классифицируют по принципу линейной последовательности расположения генов: делеции (потеря хромосомных участков), дупликации (удвоение хромосомных участков), инверсии (перевертывание на 180° относительно нормальной последовательности хромосомных участков), инсерции (вставки хромосомных участков) и транслокации (изменение расположения хромосомных участков). Подробная информация по возможным аномалиям хромосом человека представлена в главе 3.5.
Что такое конденсация хромосом
• При подготовке к расхождению хромосомы конденсируются и мигрируют к центру митотического веретена
• Хромосомы прикрепляются к микротрубочкам, выходящим из противоположных полюсов веретена, с помощью специальных участков, называемых кинетохорами
• Разрываются связи, соединяющие сестринские хроматиды вместе, и происходит их разделение
• Происходит расхождение изолированных сестринских хроматид, что предшествует цитокинезу
Как отмечалось выше, у большинства клеток одним из характерных признаков вступления в митоз является конденсация хромосом. Генетический материал интерфазных клеток упакован не так плотно как в митозе, и в процессе сегрегации хромосом важную роль играет его реорганизация с образованием компактных структур высшего порядка. Как достигается эта компактизация и каким образом она регулируется?
В конденсации хромосом участвует мультибелковый комплекс, который называется конденсин. Этот комплекс состоит из двух суперспирализованных белковых молекул, содержащих АТФазные домены и, в силу исторических причин, относящихся к семейству SMC-белков. Предполагается, что субъединицы конденсина могут «прогуливаться» по ДНК, скрепляя вместе ее различные участки. В состав конденсинового комплекса также входят другие белки, которые, наряду с SMC белками, вызывают в ДНК конформационные и топологические изменения, приводящие к компактизации.
О процессе конденсации хромосом также говорят и другие изменения связанных с ними белков, например фосфорилирование гистонов Н1 и Н3. Известно, что фосфорилирование гистона Н3 происходит с участием киназ Aurora. Фактически, фосфорилирование этого гистона в митозе обычно служит показателем активности киназы Aurora.
Образование связей между сестринскими хроматидами представляет собой процесс, родственный конденсации хромосом. При репликации в S-фазе каждая пара хроматид связывается между собой и находится в связанном состоянии до момента разделения при переходе от метафазы к анафазе. Каким образом и когда устанавливаются связи между хроматидами и как они разрываются в анафазе?
Так же как и конденсиновый комплекс, когезиновый комплекс формирует и поддерживает связи между сестринскими хроматидами, начиная от момента репликации ДНК и до начала их сегрегации. Этот комплекс состоит из двух SMC-белков, которые напоминают SMC-белки конденсина, хотя и отличаются от них, и трех дополнительных белков, Scc1, Scc3 и Pds5, также необходимых для его функционирования.
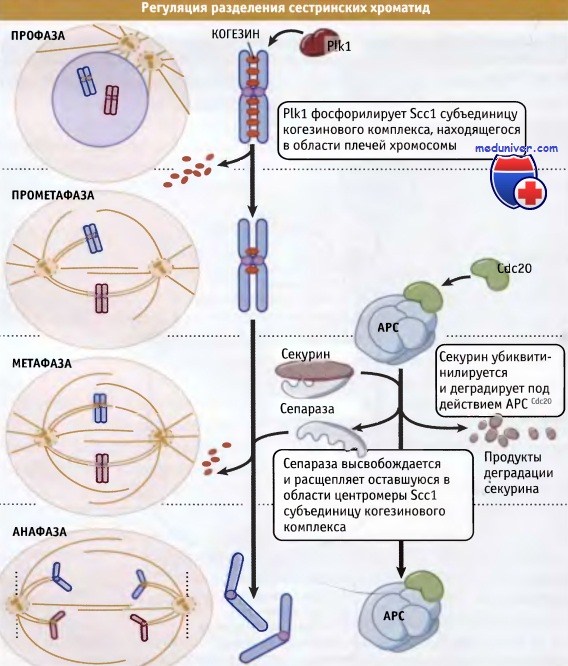 До наступления метафазы одна из протеаз, под названием сепараза, находится в связанном с секурином состоянии и не проявляет активности.
До наступления метафазы одна из протеаз, под названием сепараза, находится в связанном с секурином состоянии и не проявляет активности.
В метафазе, при действии АРС, секурин подвергается протеолизу, и таким образом происходит деблокирование сепаразы.
После этого активная сепараза расщепляет когезин, что вызывает разделение сестринских хроматид.
Предполагалось, что когезиновый комплекс скрепляет сестринские хроматиды наподобие застежки «молния». Однако недавно было показано, что SMC-белки и субъединица Scc1 обвивают сестринские хроматиды, скрепляя их. Подробности механизма этого скрепления сейчас исследуются.
Предложено два механизма разрушения связей между хроматидами. Согласно одному механизму, в профазе, с плечей хромосомы удаляется большая часть когезина, а связанный с центромерой комплекс остается на месте. Этот первый этап удаления когезина запускается Plk1, которая фосфорилирует Scc1. В то же время связанная с центромерой Scc1 когезиновая субъединица подвергается протеолизу. Если модель скрепления хроматид за счет их обвивания субъединицей Scc1 правильна, то расщепление этой субъединицы должно приводить к раскрытию обвивающего кольца и к физическому разделению сестринских хроматид. Во всяком случае, каким бы ни был механизм скрепления хроматид, очевидно, что разрушение Scc1 вызывает их диссоциацию и обеспечивает наступление анафазы.
Сепараза представляет собой сайт-специфическую протеазу, которая расщепляет Scc1 и вызывает разделение хроматид. В течение большей части клеточного цикла, сепараза поддерживается в неактивном состоянии, находясь в комплексе с другим белком, секурином. При переходе из метафазы в анафазу секурин подвергается убиквитинилированию и атакуется протеазами. При разрушении секурина высвобождается сепараза (также известная под названием сепарин), которая разрушает Scc1.
Процессы, происходящие при разделении хроматид, представлены на рисунке ниже. Таким образом, в этой простой модели ключевым процессом, запускающим сегрегацию хромосом, является протеолиз секурина. В связи с этим возникает интересный вопрос: каким образом регулируется деградация секурина?
Секурин отбирается для деградации при его взаимодействии с ЕЗ-убиквитинлигазой, известной под названием анафазный промоторный комплекс (АРС). Этот комплекс узнает белки, содержащие короткие последовательности, называемые деструктивными и/или KEN доменами (D boxes или KEN boxes). При добавлении этих последовательностей, содержащих около девяти аминокислот, в структуру белков, последние проибретают способность к деградации под действием АРС.
В отличие от SCF, для того чтобы субстрат был узнан АРС, не требуется его предварительного фосфорилирования. У почкующихся дрожжей существенная функция АРС состоит в деградации циклинов и секурина, хотя для этого комплекса известно также много других мишеней.
АРС проявляет активность только в митозе и G1-периоде, когда он предотвращает накопление циклинов и секурина. Как регулируется активность АРС? Во-первых, связывание адаптерных белков (Cdc20 и Cdh1 — обозначаемых как APC cdc20 APC cdh1 ) с APC обеспечивает значительную степень субстратной специфичности комплекса. Адаптерные белки способны связываться с АРС лишь в митозе и G1-периоде цикла. Во-вторых, активность АРС регулируется при специфическом фосфорилировании некоторых из его 13 субъединиц и адаптерных белков.
В фосфорилировании и активации АРС участвуют Cdkl и Plk1. В-третьих, активность АРС может контролироваться сигнальной системой, которая следит за прикреплением хромосом, а также точкой, контролирующей сборку веретена (SAC). Столь многочисленные уровни регуляции гарантируют, что АРС активируется только в митозе и запускает протеолиз секурина и циклина, активирует сепаразу, расщепляет когезин, и, наконец, вызывает разделение сестринских хроматид. В G1-периоде АРС сохраняет свою активность с тем, чтобы поддерживать активность Cdk1 на низком уровне. Низкая активность этой киназы позволяет сформироваться pre-RC, необходимому для следующего раунда репликации ДНК. Схема процессов активации АРС представлена на рисунке ниже.
Когда после разрыва связи сестринские хроматиды отделились друг от друга, они расходятся к противоположным полюсам клетки под действием микротрубочек веретена. Когда хромосомы разошлись и активность Cdk1 снизилась, наступает цитокинез.
Подводя итоги изложенному выше, подчеркнем, что конденсация хромосом происходит с участием белкового комплекса, который называется конденсин, и что сестринские хроматиды удерживаются вместе когезиновым комплексом до тех пор, пока не произойдет их разделение в анафазе. Разрыв связей между хроматидами требует расщепления когезина под действием протеазы, сепаразы. Для активации сепаразы необходим протеолиз ее ингибитора, секурина, который происходит с участием АРС по убиквитиновому механизму.
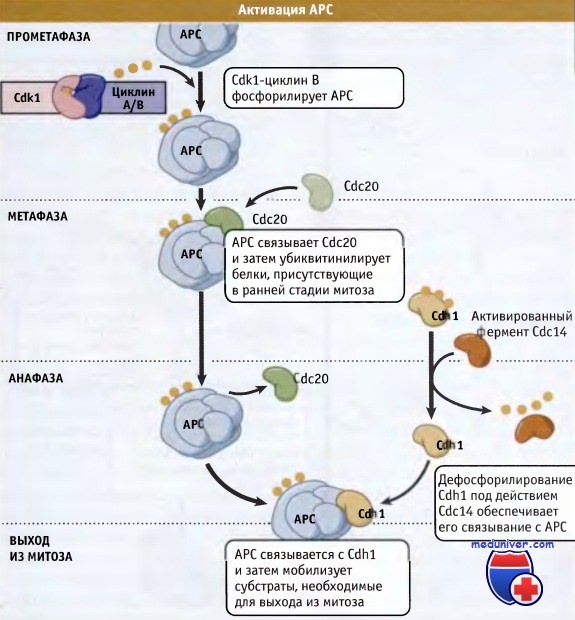 В интерфазе АРС убиквитин лигаза неактивна.
В интерфазе АРС убиквитин лигаза неактивна.
При наступлении митоза Cdk1 фосфорилирует АРС и он связывается с активатором, Cdc20.
Затем Cdc20 АРС узнает субстраты, например секурин, обеспечивая необратимое прохождение клетки по фазам митоза.
Позже в митозе АРС связывается с активатором Cdh1 и мобилизует другие субстраты, необходимые для выхода из митоза.  Первый видеокадр, показывающий хромосомы в начальных стадиях митоза.
Первый видеокадр, показывающий хромосомы в начальных стадиях митоза.
Видео процесс и фазы митоза
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Научная электронная библиотека
Юров И. Ю., Ворсанова С. Г., Воинова В. Ю., Чурносов М. И., Юров Ю. Б.,
3.1. Геном человека и структурная организация хромосом
Как уже сказано выше, данные о структуре, размерах и количестве митотических хромосом в клеточном ядре определяются как кариотип. Морфологическое строение хромосом на всех уровнях организации (молекулярном, микроскопическом и субмикроскопическом) определяется упаковкой нитей ДНК, организующих хромосому. На микроскопическом уровне нити ДНК формируют спирализованные хромонемы митотических хромосом. Упаковка хромонем по длине хромосом неравномерна. В них закономерно дифференцируются участки с резко выраженной и резко сниженной степенью спирализации хромонем. Последние формируют хромосомные перетяжки. Важным элементом структуры хромосом является первичная перетяжка, в участке которой расположена центромера. Она делит хромосому на два плеча – короткое (р) и длинное (q).
Структура хромосом претерпевает значительные изменения в ходе клеточного деления (клеточного цикла) и, следовательно, не является постоянной. Хромосомы интерфазных ядер (интерфазные хромосомы) в отличие от метафазных хромосом (см раздел, посвященный делению клетки) представляют собой более расправленные и диффузные клеточные структуры. Интерфазные хромосомы содержат одну хроматиду, в составе которой имеется одна двуспиральная молекула ДНК, в то время как метафазные хромосомы образованы двумя хроматидами и двумя молекулами ДНК. Хромосома, как функционирующая клеточная органелла, должна содержать минимум три типа последовательностей ДНК, формирующих её структурные компоненты: центромеру, теломеры и участок начала репликации ДНК.
Индивидуальные хромосомы содержат одну протяжённую двуспиральную молекулу ДНК диаметром 2 нм, которая претерпевает несколько уровней упаковки, формируя хроматин или ДНК-белковый комплекс (дезоксирибонуклеопротеид – ДНП). Элементарной дискретной единицей упаковки хроматина является нуклеосома. Каждая нуклеосома имеет вид глобулы и состоит из четырех пар гистоновых белков Н2А, Н2В, Н3 и Н4, содержащих по 102–135 аминокислотных остатков. Вокруг гистоновой глобулы диаметром 10 нм (октамера) располагается молекула двуспиральной ДНК длиной 146 пн (или 68 нм), образуя виток в 1,75 оборота. Соседние нуклеосомы соединяются так называемой «спейсерной» ДНК длиной около 20 нм, образуя структуры в виде «бус на нитке». На следующем этапе компактизации 10-нанометровые нуклеосомы образуют фибриллы хроматина диаметром 30 нм (в каждом витке примерно 6 нуклеосом), которые образуют большие петли. На стадии метафазы степень компактизации достигает 1 на 10000 от первоначальной длины хромосомной ДНК. Петли 30-нанометровых фибрилл хроматина содержат 20–100 тысяч пн. ДНК на каждой петле прикрепляется к осевой структуре хромосомы (рис. 4). Осевые компоненты метафазной хромосомы состоят из негистоновых белков.
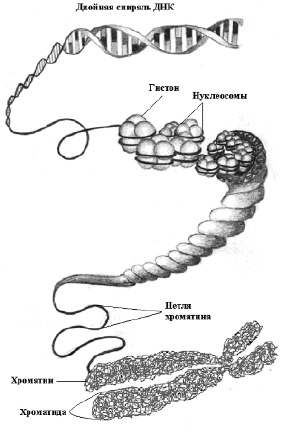
Рис. 4. Организация ДНК
Как уже отмечалось, одним из структурных элементов хромосомы является центромера – место присоединения кинетохоров (макромолекулярных структур на белковой основе, участвующих в процессе расхождения хромосом при митотическом делении клетки), и слипания сестринских хроматид. Они также регулируют присоединение хромосом к веретену деления и направляют движение хромосом в ходе мейотического и митотического деления клетки. Хромосомы без центромеры (ацентрические фрагменты) не могут присоединяться к нитям веретена деления и теряются в ходе клеточного деления. У человека центромеры содержат особые типы повторяющихся ДНК, а также специфические «центромерные» белки, обеспечивающие сегрегацию хромосом в ходе митотического и мейотического деления клеток. В зависимости от положения центромеры в хромосоме, как уже сказано выше, она может быть метацентрической, субметацентрической и акроцентрической. В акроцентрических хромосомах довольно часто на коротких плечах видны небольшого размера хроматические тельца, так называемые спутники, и спутничные нити (рис. 5). Они содержат ядрышковый организатор – значимую структуру генома человека. Плечи некоторых хромосом человека содержат вторичные перетяжки.
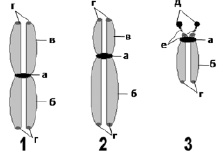
Рис. 5. Схематическое изображение метафазных хромосом:
1 – метацентрическая хромосома; 2 – субметацентрическая хромосома; 3 – акроцентрическая хромосома. Видны: а – центромеры; б – длинные плечи (q); в – короткие плечи (p); г – теломерные участки; д – спутники; е – спутничные нити
Другими структурными элементами хромосом являются теломеры. Это специализированные структуры, содержащие особые типы ДНК и белки, которые образуют концевые участки хромосом. Теломеры выполняют несколько функций:
1) поддержание структурной целостности хромосомы;
2) обеспечение полной репликации концевых участков хромосомы;
3) поддержание организации хромосом в интерфазном ядре.
Теломеры хромосом человека представляют собой повторы нуклеотидной последовательности ТТАГГГ, общий размер которой варьирует от 3 до 20 тысяч пн. Стабильность теломер поддерживается с помощью фермента теломеразы, нарушение функциональной активности которой приводит к нарушению структуры хромосом и, как следствие, к клеточной гибели. Укорочение последовательностей теломер, которому противодействует теломераза, связано с процессами старения и малигнизации.
ДНК хромосом реплицируется в ходе периода синтеза ДНК клеточного цикла. Каждая хромосома содержит множество элементарных единиц репликации – репликонов, представляющих собой участки автономной репликации. Каждый репликон имеет одну точку инициации репликации, с которой начинается двунаправленный синтез ДНК (см предыдущий раздел, посвященный синтезу ДНК). Размеры репликонов могут различаться друг от друга. Репликоны у человека могут иметь большие размеры, достигая более чем 1 млн пн.



