Физиология человека и животных
Разделы
3. Спинальные соматические и вегетативные рефлексы. Сегментарный и межсегментарный принцип работы спинного мозга
Рефлексы спинного мозга делятся на вегетативные и двигательные.
К вегетативным рефлексам относятся мочеиспускательный, дефекационный, потоотделительный, сосудистые рефлексы и др. К двигательным – кожно-мышечные, проприоцептивные и висцеромоторные рефлексы. По форме это в основном сгибательные и разгибательные рефлексы сегментарного характера.
Сгибательные и разгибательные рефлексы делятся на фазные и тонические. Фазные рефлексы – это однократное сгибание или разгибание конечности при однократном раздражении кожи или проприоцепторов. Сгибательные фазные релексы – это локтевой, подошвенный, ахиллов и т.д., разгибательный фазный рефлекс – это коленный (рисунок 2). Тонические сгибательные и разгибательные рефлексы возникают при длительном сокращении или расслаблении мышц и направлены на поддержание позы.

Рисунок 2 – Двухнейронная моносинаптическая
рефлекторная дуга коленного рефлекса
Рефлексы позы способствуют перераспределению мышечного тонуса при изменении положения тела или отдельных его частей. На уровне спинного мозга замыкаются шейные позные рефлексы, возникающие при наклоне головы или повороте головы. Эти рефлексы больше выражены у животных. Например, у кошки при наклоне головы вниз (кпереди) увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей. При наклоне головы вверх – наоборот. У человека эти рефлексы выражаются только в перераспределении мышечного тонуса для сохранения естественной позы тела. Вторая группа этих рефлексов возникает при поворотах головы вправо или влево. При этом увеличивается тонус мышц-разгибателей обеих конечностей на той стороне, куда повернута голова, и увеличивается тонус мышц-разгибателей конечностей на противоположной стороне тела. Эти рефлексы направлены на поддержание положения центра тяжести.
Универсальный принцип рефлекторной деятельности спинного мозга – это принцип общего конечного пути, открытый Ч.Шеррингтоном («воронка» Ч.Шеррингтона). Дело в том, что соотношение количества волокон в афферентных (задних корешках) и эфферентных (передних корешках) путях спинного мозга составляет примерно 5:1, поэтому широкую часть «воронки» составляют афферентные пути задних корешков, а узкую – эфферентные пути передних корешков спинного мозга.
Спинальный шок – это изменение мышечного тонуса и отсутствие рефлексов, наблюдающееся сразу после повреждения спинного мозга и связей его с вышележащими структурами ЦНС. Он развивается вследствие выключения влияния вышележащих отделов ЦНС на спинной мозг. Спинальный шок у лягушек длится несколько минут, у собак – несколько дней, у человека – около 2 месяцев. После исчезновения тормозного влияния вышележащих структур ЦНС резко увеличивается тонус мышц, иннервируемых с сегментов спинного мозга ниже места повреждения. Сгибательные и разгибательные рефлексы нижних конечностей резко повышаются. Это происходит вследствие повышения спонтанной активности γ-мотонейронов, приводящей к повышению спонтанной активности мышечных рецепторов и соответственно к повышению тонуса скелетных мышц.
Соматические рефлексы спинного мозга это
На уровне спинного мозга осуществляются многие типы сегментарных вегетативных рефлексов, большинство которых обсуждаются в других статьях на сайте (просим вас пользоваться формой поиска выше). К ним относят:
(1) изменения сосудистого тонуса в результате локального нагревания кожи;
(2) потоотделение в результате локального нагревания поверхности тела;
(3) кишечно-кишечные рефлексы, контролирующие некоторые моторные функции кишечника;
(4) брюшинно-кишечные рефлексы, тормозящие моторную активность желудочно-кишечного тракта в ответ на раздражение брюшины;
(5) эвакуационные рефлексы для опорожнения полного мочевого пузыря или переполненной толстой кишки.
Кроме того, все сегментарные рефлексы могут иногда возбуждаться одновременно в форме так называемого массивного рефлекса, изложенного далее.
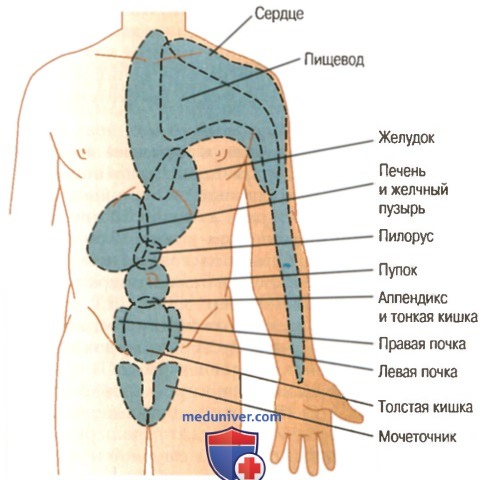 Участки поверхности тела, где появляется отраженная боль от разных внутренних органов
Участки поверхности тела, где появляется отраженная боль от разных внутренних органов 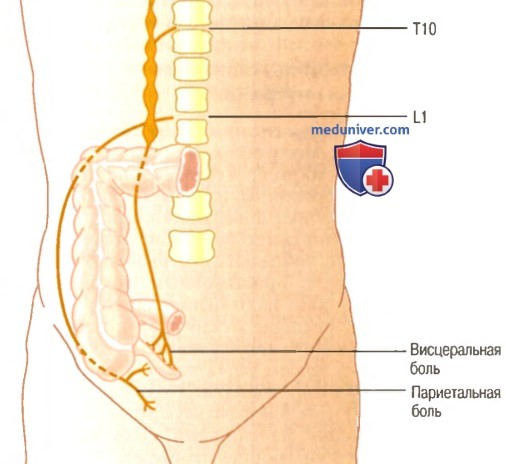 Висцеральная и париетальная передача болевых импульсов от аппендикса
Висцеральная и париетальная передача болевых импульсов от аппендикса
а) Массивный рефлекс. Иногда у спинального животного или человека активность спинного мозга чрезвычайно повышается, что сопровождается массивным импульсным разрядом в значительной его части. Обычно это происходит в связи с действием сильного болевого раздражения кожи или чрезмерного переполнения внутренних органов, например при перерастяжении мочевого пузыря или кишки. Независимо от типа стимула возникающий в результате рефлекс, называемый массивным рефлексом, вовлекает большую часть или даже весь спинной мозг.
Эффекты представляют собой:
(1) мощный сгибательный спазм значительной части скелетных мышц тела;
(2) опорожнение прямой кишки и мочевого пузыря;
(3) часто подъем артериального давления до максимальных величин, иногда до уровней систолического давления, значительно превышающих 200 мм рт. ст.;
(4) обильное потоотделение обширных областей поверхности тела.
Поскольку массивный рефлекс может продолжаться в течение нескольких минут, он, вероятно, является результатом активации большого числа реверберирующих контуров, что одновременно возбуждает большие области спинного мозга. Это похоже на механизм развития эпилептических судорог, связанный с реверберацией возбуждения, возникающей в головном мозге вместо спинного.
Пересечение спинного мозга и спинальный шок
Когда спинной мозг внезапно пересекается в верхней части шеи, сначала практически все функции спинного мозга, включая спинномозговые рефлексы, мгновенно подавляются, вплоть до полного их выключения. Эту реакцию называют спинальным шоком. Причиной такой реакции является то, что нормальная активность спинальных нейронов зависит в большой степени от постоянного тонического возбуждения спинного мозга под действием импульсов, поступающих к нему по нисходящим нервным волокнам от высших центров, особенно по ретикулоспинальным, вестибулоспинальным и кортикоспинальным трактам.
В течение нескольких часов или недель возбудимость спинальных нейронов постепенно восстанавливается. По-видимому, это общее естественное свойство нейронов повсюду в нервной системе, т.е. после потери источника облегчающих импульсов нейроны увеличивают собственную естественную степень возбудимости, чтобы компенсировать потерю хотя бы частично. Большинству неприматов, чтобы возвратить к норме возбудимость центров спинного мозга, требуется от нескольких часов до нескольких суток. Однако у человека восстановление часто задерживается на несколько недель, а иногда полного восстановления совсем не происходит. В других случаях, наоборот, происходит чрезмерное восстановление с результирующей повышенной возбудимостью некоторых или всех функций спинного мозга.
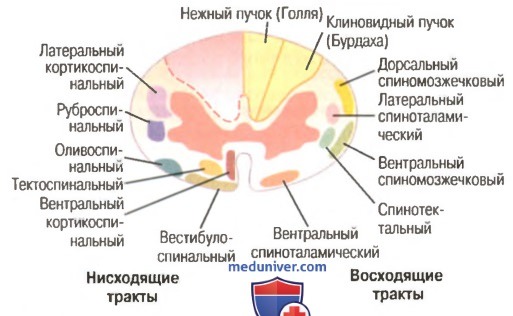 Поперечное сечение спинного мозга. Справа показаны главные восходящие тракты, слева — главные нисходящие тракты
Поперечное сечение спинного мозга. Справа показаны главные восходящие тракты, слева — главные нисходящие тракты
Далее перечислены некоторые из спинальных функций, особенно страдающих во время или после спинального шока.
1. В начале спинального шока сразу и очень значительно падает артериальное давление, иногда опускаясь ниже 40 мм рт. ст., что свидетельствует о практически полной блокаде активности симпатической нервной системы. Давление обычно возвращается к норме в течение нескольких дней (даже у человека).
2. Все рефлексы скелетных мышц, интегрированные в спинном мозге, во время первых стадий шока блокируются. Животным для восстановления этих рефлексов до нормы требуется от нескольких часов до нескольких дней; людям — от 2 нед до нескольких месяцев. И у животных, и у человека некоторые рефлексы могут в итоге стать чрезмерно возбудимыми, особенно в тех случаях, когда на фоне пересечения основной части путей между головным и спинным мозгом некоторые облегчающие пути сохраняются. Первыми восстанавливаются рефлексы на растяжение, в дальнейшем постепенно восстанавливаются более сложные рефлексы в соответствующем порядке: сгибательные, антигравитационные позные и частично шагательные.
3. Рефлексы крестцового отдела спинного мозга, контролирующие опорожнение мочевого пузыря и прямой кишки, подавлены у человека в течение первых недель после пересечения спинного мозга, но в большинстве случаев они в итоге восстанавливаются.
Видео анатомия восходящих трактов спинного мозга (восходящих путей)
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Лекция 7
Двигательные системы
Система управления движениями организована по следующему (иерархическому) принципу:
(А) Нижний уровень – спинной мозг, двигательные центры которого осуществляют рефлексы. Каждый рефлекс – это часть какого-либо движения.
(В) Высший уровень – кора больших полушарий, двигательные центры которой участвуют в осуществлении целенаправленного поведения (разнообразные движения, которые выполняются параллельно в определенной последовательности для достижения полезного результата: например, зайти в аудиторию,поздороваться, сесть, слушать лектора, записывать лекцию и т.д.).
Кроме того, в головном мозге имеются структуры, которые осуществляют контроль за выполнением движений и коррекцию (исправление) выполнения движений, используя принцип обратной связи. Таких структур две: (1) мозжечок и (2) стриопаллидарная система.
КЛАССИФИКАЦИЯ ДВИЖЕНИЙ: (1) позные (главным образом, тонические, такие как сидение, стояние и т.п.) и (2) локомоторные (главным образом, фазные, такие как ходьба, бег, плаванье, ползанье и т.д.). У человека к фазным движениям относятся также тонкие и точные движения пальцев рук (манипуляции) и речь (артикуляция).
Спинной мозг
Функции – рефлекторная и проводниковая. Рефлексы спинного мозга: (1) соматические, (2)вегетативные (симпатические и парасимпатические)
Соматические рефлексы спинного мозга
тонические: фазические (фазные):
(1) миотатический рефлекс (1) сгибательные рефлексы
(2) рефлекс с сухожильных (2) разгибательные рефлексы
рецепторов Гольджи (3) ритмические рефлексы:
(3) шейные тонические рефлексы а) чесательный
Магнуса б) шагательный
Миотатический рефлекс – рефлекторное сокращение мышцы в ответ на ее растяжение. Необходим для поддержания постоянной длины скелетной мышцы. Является частью тонического (позного) движения. Рефлекс проприоцептивный. Он возникает при растяжении проприорецепторов типа «мышечное веретено» (интрафузальное мышечное волокно), которые расположены в мышце параллельно сократительным (экстрафузальным) мышечным волокнам. Интрафузальное мышечное волокно имеет периферические – сократительные – части (типичные для поперечно-полосатой мышцы) и центральную часть – соединительнотканную сумку, где разветвляется чувствительное нервное окончание. Рефлекторная дуга моносинаптическая (двухнейронная). Схема рефлекторной дуги: при растяжении мышцы импульсы от рецепторов поступают в спинной мозг по волокнам чувствительных клеток и переключаются непосредственно на альфа-мотонейроны, иннервирующие эту скелетную мышцу. Мышца сокращается. Для того, чтобы сокращение мышцы было постоянным (поддержание позы), существует специальный механизм – гамма-эфферентная иннервация. Гамма-мотонейроны спинного мозга иннервируют проприорецептор, а именно – периферические части интрафузальных мышечных волокон, вызывая их сокращение. При этом центральная часть рецептора растягивается и импульсы от рецептора непрерывно поступают к альфа-мотонейронам, которые в свою очередь непрерывно посылают импульсы к скелетной мышце. Так поддерживается тонус скелетных мышц. Для изменения длины мышцы (перемена позы) двигательные центры ствола головного мозга посылают сигналы (1) к гамма-мотонейронам или (2) одновременно к альфа- и гамма-мотонейронам спинного мозга.
Рефлекс с сухожильных рецепторов Гольджи – рефлекторное расслабление мышцы в ответ на раздражение сухожильных рецепторов. Необходим для поддержания постоянного напряжения скелетной мышцы. Является частью тонического (позного) движения. Рефлекс проприоцептивный. Он возникает при растяжении проприорецепторов Гольджи, которые расположены последовательно в системе «мышца – сухожилие – кость» и возбуждаются при сокращении скелетной мышцы. Рефлекторная дуга полисинаптическая (трехнейронная). Схема рефлекторной дуги: при сокращении мышцы импульсы от рецепторов Гольджи поступают в спинной мозг по волокнам чувствительных клеток и переключаются на вставочные нейроны – тормозные клетки Реншоу. Клетки Реншоу тормозят альфа-мотонейроны скелетной мышцы – и мышца расслабляются (прямое постсинаптическое торможение). Примечание: блокаторы постсинаптического торможения (например, стрихнин или токсин столбнячной палочки) выключают этот тормозной рефлекс, поэтому сокращение скелетных мышц становится длительным и очень сильным, так что могут ломаться тркбчатые кости конечностей. Более того, может возникнуть асфиксия из-за длительного тетанического сокращения дыхательных мышц (нарушение вентиляции легких).
Сгибательный рефлекс –рефлекторное сокращение сгибательных мышц (отдергивание) в ответ на сильное (часто болевое) раздражение рецепторов кожи (или внутренних органов). Является частью защитного движения (например, отдергивание конечности от горячего предмета; напряжение мышц живота при раздражении брюшины). Рефлекс экстероцептивный (или интероцептивный). Рефлекторная дуга полисинаптическая (трех- или четырехнейронная). В центральной части рефлекторной дуги происходит (а) дивергенция возбуждения к нейронам мышц-синергистов, (б) реципрокное торможение нейронов мышц-антагонистов и (в) длительное последействие (несколько секунд). Происходит также распространение возбуждения к двигательным центрам противоположной конечности.
Перекрестный разгибательный рефлекс – на уровне спинного мозга сгибание одной конечности всегда сопровождается разгибанием другой конечности (через 0.2-0.5 сек). Распространение возбуждения к двигательным центрам противоположной стороны сегмента спинного мозга приводит к возбуждению мотонейронов мышц-разгибателей и реципрокному торможению мотонейронов мышц-сгибателей противоположной конечности. Перекрестный разгибательный рефлекс является частю локомоторного движения (ходьба).
Разгибательный толчок – сильное давление (удар) на подошву ноги (или ладонь руки) вызывает быстрое и сильное разгибание конечности (так называемый рефлекс опоры). Разгибательный толчок является частью тонического движения (стояние), локомоторного движения (бег, прыжок), а также защитного движения (опора на конечность предохраняет от падения и травмы тела, головы).
Чесательный рефлекс – ритмическое движение конечности (сгибание-разгибание, приведение-отведение) в ответ на слабое, движущееся раздражение кожных рецепторов. Является сложным цепным рефлексом: результат 1-го рефлекса (сгибание конечности и касание участка кожи, где действует раздражитель) является пусковым сигналом для возникновения 2-го рефлекса (разгибание и отведение конечности от участка кожи, где действует раздражитель) и т.д.
Шагательный рефлекс – шагательный автоматизм: раздражение подошвы одной конечности у спинального животного, помещенного в специальный станок, вызывает ритмические координированные шагательные движения, в которых участвуют все четыре конечности. Во-первых, каждая конечность совершает последовательное сгибание и разгибание, т.е. «шагает». Во-вторых, каждая пара конечностей (пара передних и пара задних конечностей) благодаря перекрестному разгибанию «шагают» координированно: пока левая конечность сгибается и выносится вперед, правая конечность разгибается и остается сзади, а потом наоборот.
В-третьих, шагательные движения четырех конечностей координированы по диагонали: сгибаются и выносятся вперед левая передняя и правая задняя конечности, в то время как правая передняя и левая задняя разгибаются и остаются сзади, а потом наоборот. (У человека таким образом координируются шагательные движения ног и вспомогательные движения рук при ходьбе). Итак, в спинном мозге существует врожденный нейронный контур, циркуляция возбуждения в котором приводит к выполнению жестко запрограммированного шагательного автоматизма.
Шейные тонические рефлексы Магнуса – рефлекторное перераспределение тонуса мышц конечностей при поворотах и наклонах головы. Это – наиболее сложные проявления двигательной активности спинного мозга, связанные с особым значением шейных сегментов. При поворотах и наклонах головы происходит раздражение проприорецепторов мышц шеи. Импульсы от проприорецепторов этих мышц поступают в шейные сегменты спинного мозга, которые оказывают командное влияние на двигательные центры передних конечностей (грудные сегменты) и на двигательные центры задних конечностей (поясничные сегменты спинного мозга). Например, при наклоне головы вперед у спинального животного происходит увеличение тонуса мышц-разгибателей задних конечностей и уменьшение тонуса мышц-разгибателей передних конечностей (Примечание: сгибания или разгибания конечностей у спинального животного не происходит – только изменение тонуса!).
СПИНАЛЬНОЕ ЖИВОТНОЕ – подопытное животное с перерезкой спинного мозга на высоком уровне, подготовленное для изучения спинномозговых рефлексов. У млекопитающих уровень перерезки должен быть ниже шейных сегментов, в которых находятся ядра диафрагмальных нервов (С3-5). В этом случае у спинального животного сохраняется самостоятельное дыхание.
СПИНАЛЬНЫЙ ШОК. Сразу после перерезки спинного мозга наблюдаются явления спинального шока, т.е. арефлексия – полное отсутсвие рефлексов, центры которых находятся ниже уровня перерезки. Затем рефлексы спинного мозга восстанавливаются. Продолжительность спинального шока у лягушек – меньше минуты, у птиц – несколько часов, у низших млекопитающих – несколько дней, у приматов и человека – от нескольких недель до нескольких месяцев (причем полного востановления функций спинного мозга не происходит).
Причина арефлексии – прекращение постоянных тонизирующих влияний, которые головной мозг оказывает на спинномозговые центры (по нисходящим ретикулоспинальным, вестибулоспинвльным, кортикоспинальным путям). Клиническая картина спинального шока у человека: (1) Падение артериального давления до 40 мм рт.ст. (восстанавливается до нормы в течение нескольких дней), (2) Паралич скелетной мускулатуры и отсутсвие соматических рефлексов (от 2-х недель до нескольких месяцев). Первыми появляются миотатические рефлексы, затем сгибательные, шейные тонические и шагательные рефлексы. (3) Отсутствие парасимпатических рефлексов мочеиспускания и дефекации в течение первых нескольких недель.
Рефлексы спинного мозга.
Все рефлексы спинного мозга делятся на соматические (двигательные) и вегетативные. Соматическиерефлексы делятся насухожильные(миотатические) икожные.Сухожильныерефлексывозникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению рецепторов растяжения, затем сигналы от альфа-мотонейронов спинного мозга поступают к мышцам, последние сокращаются. Характерны в первую очередь для мышц-разгибателей. В клинике определяют коленный, ахиллов, локтевой, кистевой и другие рефлексы. Коленный рефлекс имеет моносимпатический характер, т.е. в его центральной части имеется один синапс.Кожныерефлексыобусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной. Спинальные нервные центры находятся под контролем вышележащих нервных центров. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшается.
Вегетативные рефлексы спинного мозгаделятся насимпатическиеипарасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыделения, дефекации, эрекции, эякуляции и т.д.
Функции продолговатого мозга.
Основными функциями продолговатого мозга являются проводниковая,рефлекторнаяиассоциативная. Первая осуществляется проводящими путями, проходящими через него. Вторая – нервными центрами. В ромбовидной ямке продолговатого мозга находятся ядра X, XI, XII пар черепномозговых нервов, а также ретикулярная формация. Рефлекторные функции делятся на соматические и вегетативные.Соматическимиявляются статические рефлексы продолговатого мозга, относящиеся к позно-тоническим или рефлексам позы. Эти рефлексы осуществляются ядром Дейтерса из группы вестибулярных ядер. От него к мотонейронам разгибателей спинного мозга идут нисходящие вестибулоспинальные тракты. Рефлексы возникают тогда, когда возбуждаются вестибулярные рецепторы или проприорецепторы мышц шеи. Коррекция положения тела происходит за счет изменения тонуса мышц. Например, при запрокидывании головы животного назад повышается тонус разгибателей передних конечностей и снижается тонус разгибателей задних. При наклоне головы вперед возникает обратная реакция. При повороте головы в сторону, повышается тонус разгибателей конечности на этой стороне и сгибателей противоположной конечности.
В продолговатом мозге находятся жизненно важные центры. К ним относятся дыхательный,сосудодвигательныйцентрыицентррегуляциисердечнойдеятельности. Первый обеспечивает смену фаз дыхания, второй – тонус периферических сосудов, третий – регуляцию частоты и силы сердечных сокращений.
В области ядер блуждающего нерва находятся центры слюноотделения, секреции желудочных, кишечных желез, поджелудочной железы и печени. Здесь же расположеныцентры регуляции моторики пищеварительного канала.
Важной функцией продолговатого мозга является формирование защитных рефлексов. В нем находятся рвотный цент, центры кашля, чихания, смыкания век и слезотеченияпри раздражении роговицы.
Здесь расположены бульбарные отделы центров, участвующих в организации пищевых рефлексов – сосания, жевания, глотания.
В продолговатом мозге происходит первичный анализ ряда сенсорных сигналов. В частности; в нем расположены ядра слухового нерва, верхнее вестибулярное ядро, а к ядрам языкоглоточного нерва поступают сигналы от вкусовых рецепторов. От рецепторов кожи лица они идут к ядрам тройничного нерва.
25. Соматические рефлексы спинного мозга.
Классификация соматических рефлексов спинного мозга. Все спинальные рефлексы можно объединить в две группы по следующим признакам. Во-первых, по рецепторам, раздражение которых вызывает рефлекс: а) проприорецептивные, б) висцероре цептивные и в) кожные рефлексы (защитные). Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висцерорецептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины.
Во-вторых, спинальные рефлексы целесообразно объединить по органам (эффекторам рефлекса): а) рефлексы конечностей, б) брюшные, в) органов таза. Рассмотрим каждую группу.
Рефлексы конечностей. Данная группа рефлексов наиболее обширная, в клинической практике их исследуют наиболее часто. Однако отдельные рефлексы описывают без взаимной связи. Если в качестве объединяющего признака рефлексов конечностей принять характер ответной реакции, то все их можно объединить в четыре группы: 1) сгибательные, 2) разгибательные, 3) ритмические и 4) познотонические.
Сгибательные рефлексы делятся на фазные и тонические.
Фазные рефлексы — это однократное сгибание конечности при однократном раздражении кожи или проприорецепторов. Одновременно с возбуждением мотонейронов мышц-сгибателей происходит реципрокное торможение мотонейронов мышц-разгибателей. Рефлексы, возникающие с рецепторов кожи, являются по-лисинаптическими, они имеют защитное значение. Например, погружение в слабый раствор серной кислоты лапки Финальной лягушки, подвешенной на крючке, или щипок кожи конечности пинцетом вызывают отдергивание конечности вследствие сгибания ее в коленном суставе, а при более сильном раздражении — и в тазобедренном суставе. Рефлексы, возникающие Проприорецепторов, могут быть моносинаптическими и поли-Инаптическими, например шейные познотонические рефлексы, азные рефлексы с проприорецепторов участвуют в формировали акта ходьбы. По степени выраженности фазных сгибательных или двигательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения. на сгибательные, а также разгибательные рефлексы возникают при длительном растяжении мышц, их главное назначения поддержание позы. Тоническое сокращение скелетных мышц является фоновым для осуществления всех двигательных актов, осуществляемых с помощью фазических сокращений мышц. В клинике исследуют несколько сгибательных фазных рефлексов: локтевой и ахиллов (проприоцептивные рефлексы), подошвенный (кожный рефлекс). Локтевой рефлекс выражается в сгибании рук в локтевом суставе, возникает при ударе молоточком по сухожилию (при вызове рефлекса рука должна быть слегка согнута в локтевом суставе), его дуга замыкается в пятом-шестом шейных сегментах спинного мозга. Ахиллов рефлекс выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию, рефлекторная дуга замыкается на уровне крестцовых сегментов. Подошвенный рефлекс — сгибание стопы и пальцев при штриховом раздражении подошвы, дуга рефлекса замыкается также на уровне.
Разгибательные рефлексы, как и сгибательные, бывают фазными и тоническими, возникают с проприорецепторов мышц-разгибателей, являются моносинаптическими. Одновременно со сгибательным рефлексом возникает перекрестный разгибательный рефлекс другой конечности.
Фазные рефлексы возникают в ответ на однократное раздражение мышечных рецепторов, например при ударе по сухожилию четырехглавой мышцы ниже надколенной чашечки. При этом возникает коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся — постсинаптическое реципрокное торможение с помощью вставочных тормозных клеток Реншоу). Рефлекторная дуга коленного рефлекса замыкается во втором—четвертом поясничных сегментах. Фазные разгибательные рефлексы участвуют, как и сгибательные, в формировании акта ходьбы.
Тонические разгибательные рефлексы представляют собой длительное сокращение мышц-разгибателей при длительном растяжении их сухожилий. Их роль — поддержание позы. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют также миотатическими.
Вторая группа шейных позных рефлексов возникает с тех же рецепторов, но только при поворотах головы вправо или влево. Притом повышается тонус мышц-разгибателей обеих конечностей к стороне, куда повернута голова, и повышается тонус мышц-згибателей на противоположной стороне. Рефлекс направлен на Ранение позы, которая может быть нарушена вследствие изменения положения центра тяжести после поворота головы. центр тяжести смещается в сторону поворота головы — именно на ЭТОЙ стороне повышается тонус мышц-разгибателей обеих конечностей
Ритмические рефлексы — многократное повторное сгибание и разгибание конечностей. Примерами этих рефлексов могут быть рефлексы потирания у лягушки, чесательный и шагательный рефлексы у собаки. Рефлекс потирания заключается в том, что после смазывания раствором кислоты кожи бедра спинальная лягушка (лягушка, у которой удален головной мозг) многократно потирает этот участок, т.е. пытается освободиться от раздражителя. Слабое раздражение кожи туловища у собаки вызывает почесывание этого участка задней конечностью. Чесательный рефлекс — аналог рефлекса потирания у лягушки. Шагательный рефлекс наблюдают у спинальной собаки, подвешенной с помощью лямок в станке.
Брюшные рефлексы (верхний, средний и нижний) вызываются штриховым раздражением кожи живота, выражаются в сокращении соответствующих участков мускулатуры стенки живота. Это защитные рефлексы. Для вызова верхнего брюшного рефлекса раздражение наносят параллельно нижним ребрам непосредственно под ними, дуга рефлекса замыкается на уровне грудных сегментов спинного мозга. Средний брюшной рефлекс вызывают раздражением на уровне пупка (горизонтально), дуга рефлекса замыкается на уровне. Для получения нижнего брюшного рефлекса раздражение наносят параллельно паховой складке (рядом с ней), дуга рефлекса замыкается на уровне.
26.Структурно-функциональная характерпистика ствола мозга.
Особенности функциональной организации. Продолговатый мозг (medulla oblongata) у человека имеет длину около 25 мм. Он является продолжением спинного мозга. Структурно по разнообразию и строению ядер продолговатый мозг сложнее, чем спинной. В отличие от спинного мозга он не имеет метамерного, повторяемого строения, серое вещество в нем расположено не в центре, а ядрами к периферии.
В продолговатом мозге находятся оливы, связанные со спинным мозгом, экстрапирамидной системой и мозжечком — это тонкое и клиновидное ядра проприоцептивной чувствительности (ядра Голля и Бурдаха). Здесь же находятся перекресты нисходящих пирамидных путей и восходящих путей, образованных тонким и клиновидным пучками (Голля и Бурдаха), ретикулярная формация.
Продолговатый мозг за счет своих ядерных образований и ретикулярной формации участвует в реализации вегетативных, соматических, вкусовых, слуховых, вестибулярных рефлексов. Особенностью продолговатого мозга является то, что его ядра, возбуждаясь последовательно, обеспечивают выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, что наблюдается, например, при глотании.
В продолговатом мозге расположены ядра следующих черепных нервов:
пара VIII черепных нервов — преддверно-улитковый нерв состоит из улитковой и преддверной частей. Улитковое ядро лежит в продолговатом мозге;
пара IX — языкоглоточный нерв (п. glossopharyngeus); его ядро образовано 3 частями — двигательной, чувствительной и вегетативной. Двигательная часть участвует в иннервации мышц глотки и полости рта, чувствительная — получает информацию от рецепторов вкуса задней трети языка; вегетативная иннервирует слюнные железы;
пара X — блуждающий нерв (n.vagus) имеет 3 ядра: вегетативное иннервирует гортань, пищевод, сердце, желудок, кишечник, пищеварительные железы; чувствительное получает информацию от рецепторов альвеол легких и других внутренних органов и двигательное (так называемое обоюдное) обеспечивает последовательность сокращения мышц глотки, гортани при глотании;
пара XI — добавочный нерв (n.accessorius); его ядро частично расположено в продолговатом мозге;
пара XII — подъязычный нерв (n.hypoglossus) является двигательным нервом языка, его ядро большей частью расположено в продолговатом мозге.
Сенсорные функции. Продолговатый мозг регулирует ряд сенсорных функций: рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; рецепцию слуховых раздражений — в ядре улиткового нерва; рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). На уровне продолготоватого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
Проводниковые функции. Через продолготоватый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
Рефлекторные функции. Многочисленные рефлексы продолговатого мозга делят на жизненно важные и нежизненно важные, однако такое представление достаточно условно. Дыхательные и сосудодвигательные центры продолговатого мозга можно отнести к жизненно важным центрам, так как в них замыкается ряд сердечных и дыхательных рефлексов.
Продолговатый мозг организует и реализует ряд защитных рефлексов: рвоты, чиханья, кашля, слезоотделения, смыкания век. Эти рефлексы реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга, отсюда идет команда к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного или подъязычного нервов, в результате реализуется тот или иной защитный рефлекс. Точно так же за счет последовательного включения мышечных групп головы, шеи, грудной клетки и диафрагмы организуются рефлексы пищевого поведения: сосания, жевания, глотания.
Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо.
Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы, соответствующей моменту прямолинейного или вращательного движения.
Большая часть автономных рефлексов продолговатого мозга реализуется через расположенные в нем ядра блуждающего нерва, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции названных органов.
Возбуждение ядер блуждающего нерва вызывает усиление сокращения гладких мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов. При этом замедляется и ослабляется работа сердца, сужается просвет бронхов.
Деятельность ядер блуждающего нерва проявляется также в усилении секреции бронхиальных, желудочных, кишечных желез, в возбуждении поджелудочной железы, секреторных клеток печени.
В продолговатом мозге локализуется центр слюноотделения, парасимпатическая часть которого обеспечивает усиление общей секреции, а симпатическая — белковой секреции слюнных желез.
В структуре ретикулярной формации продолговатого мозга расположены дыхательный и сосудодвигательный центры. Особенность этих центров в том, что их нейроны способны возбуждаться рефлекторно и под действием химических раздражителей.
Дыхательный центр локализуется в медиальной части ретикулярной формации каждой симметричной половины продолговатого мозга и разделен на две части, вдоха и выдоха.
В ретикулярной формации продолговатого мозга представлен другой жизненно важный центр — сосудодвигательный центр (регуляции сосудистого тонуса). Он функционирует совместно с вышележащими структурами мозга и прежде всего с гипоталамусом. Возбуждение сосудодвигательного центра всегда изменяет ритм дыхания, тонус бронхов, мышц кишечника, мочевого пузыря, цилиарной мышцы и др. Это обусловлено тем, что ретикулярная формация продолговатого мозга имеет синаптические связи с гипоталамусом и другими центрами.
В средних отделах ретикулярной формации находятся нейроны, образующие ретикулоспинальный путь, оказывающий тормозное влияние на мотонейроны спинного мозга. На дне IV желудочка расположены нейроны «голубого пятна». Их медиатором является норадреналин. Эти нейроны вызывают активацию ретикулоспинального пути в фазу «быстрого» сна, что приводит к торможению спинальных рефлексов и снижению мышечного тонуса.
Симптомы повреждения. Повреждение левой или правой половины продолговато мозга выше перекреста восходящих путей проприоцептивной чувствительности вызывает на стороне повреждения нарушения чувствительности и работы мышц лица и головы. В то же время на противоположной стороне относительно стороны повреждения наблюдаются нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это объясняется тем, что восходящие и нисходящие проводящие пути из спинного мозга и в спинной мозг перекрещиваются, а ядра черепных нервов иннервируют свою половину головы, т. е. черепные нервы не перекрещиваются.
Мост (pons cerebri, pons Varolii) располагается выше продолговатого мозга и выполняет сенсорные, проводниковые, двигательные, интегративные рефлекторные функции.
В состав моста входят ядра лицевого, тройничного, отводящего, преддверно-улиткового нерва (вестибулярные и улитковые ядра), ядра преддверной части преддверно-улиткового нерва (вестибулярного нерва): латеральное (Дейтерса) и верхнее (Бехтерева). Ретикулярная формация моста тесно связана с ретикулярной формацией среднего и продолговатого мозга.
Важной структурой моста является средняя ножка мозжечка. Именно она обеспечивает функциональные компенсаторные и морфологические связи коры большого мозга с полушариями мозжечка.
Сенсорные функции моста обеспечиваются ядрами преддверно-улиткового, тройничного нервов. Улитковая часть преддверно-улиткового нерва заканчивается в мозге в улитковых ядрах; преддверная часть преддверно-улиткового нерва — в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений их силы и направленности.
Чувствительное ядро тройничного нерва получает сигналы от рецепторов кожи лица, передних отделов волосистой части головы, слизистой оболочки носа и рта, зубов и конъюнктивы глазного яблока. Лицевой нерв (п. facialis) иннервирует все мимические мышцы лица. Отводящий нерв (п. abducens) иннервирует прямую латеральную мышцу, отводящую глазное яблоко кнаружи.
Двигательная порция ядра тройничного нерва (п. trigeminus) иннервирует жевательные мышцы, мышцу, натягивающую барабанную перепонку, и мышцу, натягивающую небную занавеску.
Проводящая функция моста. Обеспечивается продольно и поперечно расположенными волокнами. Поперечно расположенные волокна образуют верхний и нижний слои, а между ними проходят идущие из коры большого мозга пирамидные пути. Между поперечными волокнами расположены нейронные скопления — ядра моста. От их нейронов начинаются поперечные волокна, которые идут на противоположную сторону моста, образуя среднюю ножку мозжечка и заканчиваясь в его коре.
В покрышке моста располагаются продольно идущие пучки волокон медиальной петли (lemniscus medialis). Они пересекаются поперечно идущими волокнами трапециевидного тела (corpus trapezoideum), представляющие собой аксоны улитковой части преддверно-улиткового нерва противоположной стороны, которые заканчиваются в ядре верхней оливы (oliva superior). От этого ядра идут пути боковой петли (lemniscus lateralis), которые направляются в заднее четверохолмие среднего мозга и в медиальные коленчатые тела промежуточного мозга.
В покрышке мозга локализуются переднее и заднее ядра трапециевидного тела и латеральной петли. Эти ядра вместе с верхней оливой обеспечивают первичный анализ информации от органа слуха и затем передают информацию в задние бугры четверохолмий.
В покрышке также расположены длинный медиальный и тектоспинальный пути.
Собственные нейроны структуры моста образуют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательной порции ядра и среднее сенсорное ядро тройничного нерва.
Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга и началом этой же системы среднего мозга. Аксоны нейронов ретикулярной формации моста идут в мозжечок, в спинной мозг (ретикулоспинальный путь). Последние активируют нейроны спинного мозга.
Ретикулярная формация моста влияет на кору большого мозга, вызывая ее пробуждение или сонное состояние. В ретикулярной формации моста находятся две группы ядер, которые относятся к общему дыхательному центру. Один центр активирует центр вдоха продолговатого мозга, другой — центр выдоха. Нейроны дыхательного центра, расположенные в мосте, адаптируют работу дыхательных клеток продолговатого мозга в соответствии с меняющимся состоянием организма.



