Вопрос 36.8 Физиология среднего мозга, его рефлекторная деятельность и участие в процессах саморегуляции функций.
Средний мозг представлен четверохолмием и ножками мозга. Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и ядра черепных (глазодвигательного и блокового) нервов, а также ядра ретикулярной формации.
Средний мозг играет важную роль в регуляции мышечного тонуса и в осуществлении установочных и выпрямительных рефлексов, благодаря которым возможны стояние и ходьба.
Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора, нижние — слухового. В них происходит первичное переключение зрительной и слуховой информации. От бугров четверохолмия аксоны их нейронов идут к ретикулярной формации ствола, мотонейронам спинного мозга. Нейроны четверохолмия могут быть полимодальными и детекторными (реагируют на 1 признак раздражения). Основная функция бугров четверохолмия — организация реакции настораживания и так называемых старт-рефлексов на внезапные зрительные или звуковые сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции. Четверохолмие организует ориентировочные зрительные и слуховые рефлексы. У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, вскрикивание. При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
37.9 Роль среднего и продолговатого мозга в регуляции мышечного тонуса. Децеребрационнаярегидность и механизм ее возникновения (гамма-регидность).
Продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо.
Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы, соответствующей моменту прямолинейного или вращательного движения.
Возбуждение ядер блуждающего нерва вызывает усиление сокращения гладких мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов.
Средний мозг. Красные ядра располагаются в верхней части ножек мозга. Они связаны с корой большого мозга (нисходящие от коры пути), подкорковыми ядрами, мозжечком, спинным мозгом (красноядерно-спинномозговой путь). Базальные ганглии головного мозга, мозжечок имеют свои окончания в красных ядрах. Нарушение связей красных ядер с ретикулярной формацией продолговатого мозга ведет к децеребрационной ригидности. Это состояние характеризуется сильным напряжением мышц-разгибателей конечностей, шеи, спины. Основной причиной возникновения децеребрационной ригидности служит выраженное активирующее влияние латерального вестибулярного ядра (ядро Дейтерса) на мотонейроны разгибателей. Это влияние максимально в отсутствие тормозных влияний красного ядра и вышележащих структур, а также мозжечка. При перерезке мозга ниже ядра латерального вестибулярного нерва децеребрационная ригидность исчезает.
Красные ядра, получая информацию от двигательной зоны коры большого мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, посылают корригирующие импульсы к мотонейронам спинного мозга по руброспинальному тракту и тем самым регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению.
Другое функционально важное ядро среднего мозга — черное вещество — располагается в ножках мозга, регулирует акты жевания, глотания (их последовательность), обеспечивает точные движения пальцев кисти руки, например при письме. Нейроны этого ядра способны синтезировать медиатор дофамин, который поставляется аксональным транспортом к базальным ганглиям головного мозга. Поражение черного вещества приводит к нарушению пластического тонуса мышц. Тонкая регуляция пластического тонуса при игре на скрипке, письме, выполнении графических работ обеспечивается черным веществом.
39.Средний мозг,его роль в первичных ориентировочных рефлексах и регуляции мышечного тонуса.Децеребрационная ригидность
Средний мозг представлен четверохолмием и ножками мозга. Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и ядра черепных (глазодвигательного и блокового) нервов, а также ядра ретикулярной формации.
Средний мозг играет важную роль в регуляции мышечного тонуса и в осуществлении установочных и выпрямительных рефлексов, благодаря которым возможны стояние и ходьба.
Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора, нижние — слухового. В них происходит первичное переключение зрительной и слуховой информации. От бугров четверохолмия аксоны их нейронов идут к ретикулярной формации ствола, мотонейронам спинного мозга. Нейроны четверохолмия могут быть полимодальными и детекторными (реагируют на 1 признак раздражения). Основная функция бугров четверохолмия — организация реакции настораживания и так называемых старт-рефлексов на внезапные зрительные или звуковые сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции. Четверохолмие организует ориентировочные зрительные и слуховые рефлексы. У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, вскрикивание. При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.



40.Тонические рфлексы,их виды и значение
В связи с перераспределением мышечного тонуса у человека и позвоночных животных образовались особые тонические рефлексы. Они осуществляются в ответ на возбуждение рецепторов, находящихся в мышцах, сухожилиях, коже, глазах и органах равновесия, и вызывают деятельность различных мышц. Тонические рефлексы делятся на три группы: рефлексы позы, выпрямительные и статокинетические. Рефлексы позы проявляются в случае изменения положения тела, при котором возникает угроза потери равновесия. Такие рефлексы помогают удержать равновесие и тем самым сохранить нормальное положение тела.Рефлексы позы возникают при изменении положения головы по отношению к туловищу, т. е. при наклонах головы вперед, назад и в стороны. Наклон головы вперед вызывает увеличение напряжения мышц, сгибающих туловище. Наклон головы назад увеличивает напряжение разгибателей туловища.Выпрямительные рефлексы проявляются при нарушении вертикального положения тела. Они играют защитную роль, оберегая тело от внезапного падения, например у поскользнувшегося или оступившегося человека.
11.2. Участие среднего мозга в регуляции движений и позного тонуса
Горизонтальные движения глаза зависят от содружественной работы наружной и внутренней прямых мышц глаза. Нейрофизиологическими исследованиями установлено, что степень возбуждения мотонейронов, локализованных в ядрах отводящего и глазодвигательного нервов, идущих к этим мышцам, контролируется центрами ретикулярной формации варолиева моста. В этих центрах обнаружены нейроны, которые характеризуются повышением частоты своей импульсной активности перед началом каждой горизонтальной саккады. Другая группа нейронов, напротив, прерывает свои импульсные разряды до и во время саккад.
Очевидно, характер разрядов нейронов ретикулярного центра определяет его активирующие или, наоборот, тормозные управляющие воздействия на мотонейроны отводящего и глазодвигательного нервов в соответствии с принципом антагонистической иннервации. Аналогичным образом при вертикальных движениях глаза также наблюдается антагонистическое взаимодействие между двумя другими группами глазных мышц. При движении глаза по вертикали вверх сокращаются нижняя косая и верхняя прямая мышцы (глазодвигательный нерв) и одновременно расслабляются верхняя косая (блоковый нерв) и нижняя прямая (глазодвигательный нерв ) мышцы глаза. В данном случае возбудимость соответствующих моторных центров регулируется группой нейронов ретикулярной формации среднего мозга, локализованной под верхними холмами четверохолмия. В этой области также обнаружены ретикулярные нейроны, увеличивающие частоту своих импульсных разрядов перед началом вертикальных саккад. Поражение ретикулярной формации среднего мозга при патологических процессах парализует вертикальные движения глаз.
Таким образом, ретикулярная формация среднего мозга играет важную роль в координации сокращений глазных мышц. Она получает афферентные входы от верхних холмов четверохолмия, мозжечка, вестибулярных ядер, зрительных областей коры полушарий головного мозга. Поступающие по этим входам сигналы интегрируются центрами ретикулярной формации и служат для рефлекторного изменения работы глазодвигательного аппарата при внезапном появлении движущихся объектов, при изменении положения головы, при произвольных движениях глаз и т. д. По отношению к моторным центрам в ядрах черепно-мозговых нервов ретикулярная формация выступает как более высокий уровень регуляции глазных движений, осуществляемой за счет возбуждающих и тормозных влияний.
Одной из структур, ответственных за надсегментарный контроль позного тонуса, является красное ядро среднего мозга. Будучи составной частью организующей движения экстрапирамидной системы, красное ядро получает входы от моторной коры, ядер мозжечка, черной субстанции среднего мозга и дает начало руброспинальному тракту, который как общий путь обеспечивает регуляцию тонуса скелетных мышц.
Существует точка зрения, что красное ядро и ядро Дейтерса оказывают друг на друга тормозное воздействие, которое в норме снижает тонус антигравитационных разгибательных мышц. О наличии такого взимодействия свидетельствует классический опыт Ч. Шеррингтона с перерезкой стволовой части мозга. Если перерезку произвести на уровне переднего края задних холмов четверохолмия и таким образом отделить красное ядро от заднего мозга, где локализовано вестибулярное ядро Дейтерса, то у кошки развивается децеребрационная ригидность.
Это состояние характеризуется резким повышением тонуса мышц-разгибателей конечностей, спины и хвоста. Оперированное животное, поставленное на лапы, сохраняет стоячее положение, так как сгибания в суставах не происходит. Если плоскость перерезки сдвинуть вперед и сохранить связь красного ядра с задним мозгом, то состояние децеребрационной ригидности не возникает. Децеребрационная ригидность устраняется при коагуляции ядра Дейтерса или при повторной перерезке стволовой части мозга каудальнее зоны расположения вестибулярных ядер.Все эти факты указывают на то, что основной причиной децеребрационной ригидности является снятие при перерезке тормозных влияний со стороны красного ядра на ядро Дейтерса. При этом начинает преобладать действие ядра Дейтерса, которое, как известно, возбуждает мотонейроны мышц разгибателей. По представлениям Ч. Шеррингтона, красное ядро является не только источником, но и посредником в регуляции разгибательных рефлексов, передавая тормозные влияния со стороны мозжечка и моторных областей коры. Удаление мозжечка или моторной коры сопровождается усилением тонуса разгибательных мышц.
Физиологические основы поддержания равновесия
Причиной головокружения в большинстве случаев служит нарушение согласованной деятельности различных сенсорных систем – вестибулярной, зрительной, проприоцептивной (информация о положении тела в пространстве, получаемая от рецепторов, расположенных главным образом в мышцах и сухожилиях). Кроме того, важной, а иногда и доминирующей причиной возникновения головокружения является дисфункция центральных структур, участвующих в поддержании равновесия тела, главным образом, ядер мозжечка.
Вестибулярная система
Вестибулярная система состоит из:
Правильная работа вестибулярной системы позволяет человеку четко ориентироваться в трехмерном пространстве, а именно:
Лабиринт располагается в каменистой части височной кости и включает:
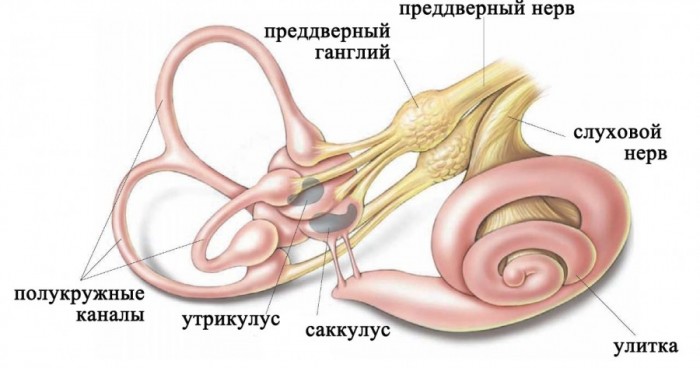
Строение лабиринта
В каждой камере отолитового аппарата и в каждом полукружном канале имеется скопление рецепторных клеток – макула, которая покрыта желатинообразной массой – купулой. В отолитовом аппарате купула покрывает волосковые клетки наподобие подушки и содержит отложения кристаллов кальцита (отолиты), которые придают купуле дополнительный вес.
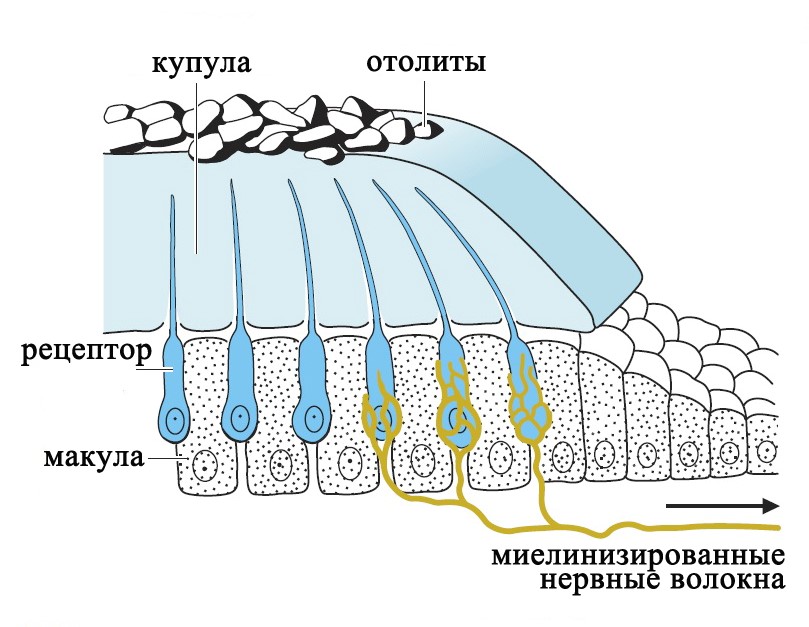
Отолитовый аппарат
В полукружных каналах желатинообразная масса не содержит отолитов и полностью перекрывает просвет канала.
Рецепторы вестибулярной системы представлены волосковыми клетками, которые несут на апикальной поверхности от 60 до 80 тонких выростов цитоплазмы (стереоцилий) и одну ресничку (киноцилию).
Восприятие положения тела относительно силы гравитации
При вертикальном положении головы макула утрикулуса располагается горизонтально. Когда голова наклоняется в сторону, утяжеленная отолитами желатинообразная мембрана под действием силы тяжести соскальзывает в сторону наклона. Это скольжение приводит к изгибанию стереоцилей волосковых клеток. Наклон стереоцилей сопровождается (в зависимости от направления) повышением или снижением частоты нервных импульсов в чувствительных нейронах вестибулярного ганглия. Макула саккулуса располагается вертикально и действует таким же образом.
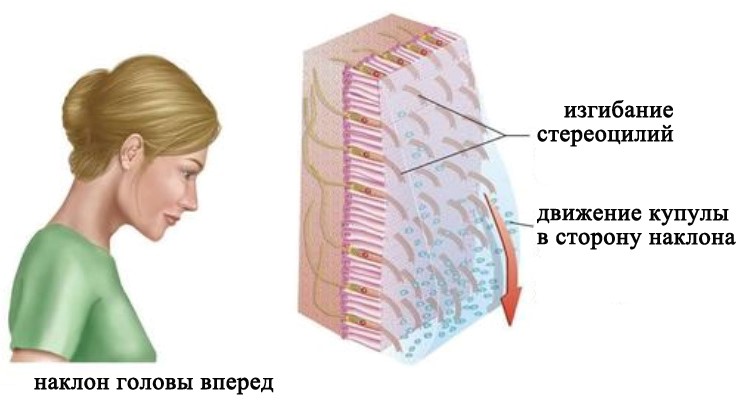
Восприятие положения тела относительно силы гравитации
Восприятие линейных ускорений
При резком линейном ускорении тела купула саккулуса или утрикулуса за счет сил инерции смещается в направлении, противоположном направлению движения, что также приводит к изменению электрической активности рецепторов.
Восприятие углового ускорения
Три полукружных канала расположены в трех разных плоскостях. Каждый из трех каналов действует как замкнутая трубка, заполненная лимфой. В расширенной части канала его внутренняя стенка выстлана волосковыми клетками, а расположенная над ними купула полностью перекрывает просвет канала. При повороте головы полукружные каналы поворачиваются вместе с ней, а эндолимфа в силу своей инерции в первый момент остается на месте. В результате этого возникает разность давлений по обе сторону купулы, и она прогибается в направлении, противоположном движению. Это вызывает деформацию стереоцилий и последующее изменение активности нейронов.
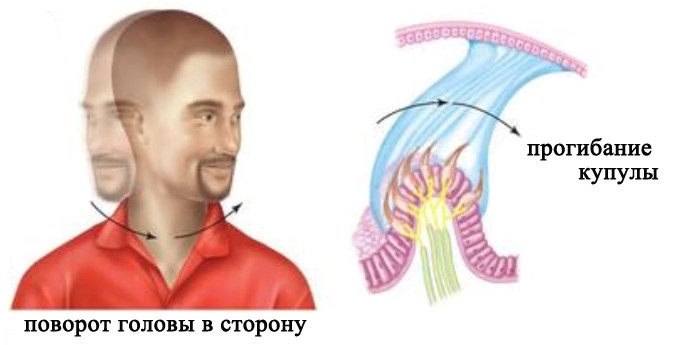
Восприятие углового ускорения
При вращении головы только в горизонтальной, сагитальной или фронтальной плоскости активируются рецепторы одного из соответствующих каналов. При сложном вращении головы активируются рецепторы всех трех каналов. Информация от них поступает в головной мозг и на основе ее конвергенции и анализа модулируется истинная картина перемещения головы.
Центральный отдел вестибулярной системы
Аксоны чувствительных нейронов, тела которых располагаются в вестибулярном ганглии, следуют в продолговатый мозг и оканчиваются в четырех парных вестибулярных ядрах. Приходящие в эти ядра импульсы от рецепторов дают точную информацию о положении в пространстве исключительно головы (но не всего тела!), поскольку она может быть наклонена или повернута относительно туловища. Для восприятия положения тела в пространстве необходим также учет угла наклона и поворота головы относительно туловища, поэтому вестибулярные ядра получают дополнительные стимулы от проприорецепторов мышц шеи.
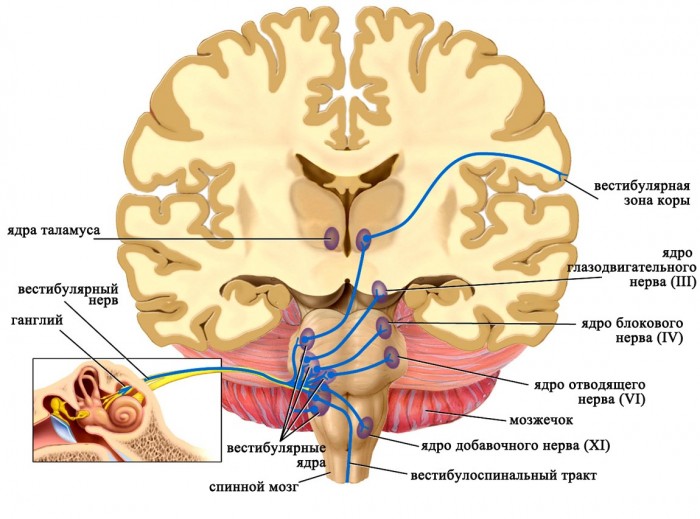 Ядра вестибулярного нерва и их связи
Ядра вестибулярного нерва и их связи
Далее от вестибулярных ядер афферентная импульсация направляется к нейронам специфических ядер таламуса, а отростки последних достигают постцентральной извилины коры больших полушарий головного мозга
Проприоцептивная система
Благодаря проприоцепции, мы ощущаем положение конечностей, движение и степень мышечного напряжения в них. Это дает человеку чувство “опоры”, т.е. осознание, что стопы опираются на какую-либо поверхность, удерживая вес тела. Рецепторный аппарат проприоцептивной чувствительности, расположен в мышцах, сухожилиях, фасциях, капсулах суставов, а также в коже.
Необходимо отметить, что важную роль в поддержании равновесия тела играют рецепторы глубокой чувствительности, расположенные не только в конечностях, но и в структурах шеи, главным образом, в глубоких мышцах. Информация, получаемая головным мозгом от этих рецепторов, необходима для пространственной ориентации человека, поддержании его позы, а также координинации движения головы и туловища.
Зрительная система
Эффективное поддержание равновесия требует четкого контроля со стороны зрительной системы (в соответствие с принципом обратной связи). При этом контроль над движениями мышц глазного яблока является чрезвычайно сложным процессом. Существует 3 основных системы контроля взора:
В пределах головного мозга эти системы контролируются определенными анатомическими зонами, которые являются в значительной степени изолированными, и обеспечивают две главные функции:
Система саккадических движений глазных яблок
Когда объект интереса появляется в периферии визуальной области, происходит быстрый поворот глазных яблок в его сторону, так, что изображение объекта проецируется на сетчатку в области желтого пятна. Тот же самый двигательный ответ глазных яблок может быть вызван внезапным звуком или болезненным стимулом. Такое быстрое движение глаз называется саккадическим, от французского слова, означающего резкое движение парусника при ветре или дергание головы лошади от потягивания узды. В целом, система саккадических движений глазных яблок обеспечивает обнаружение зрительной цели и выведение ее на наиболее чувствительную часть сетчатой оболочки. Саккады возникают, например, в процессе чтения, при этом глаза человека обычно совершают несколько саккадических движений на каждой строке. Кроме того, они появляются, когда человек рассматривает какой-либо объект (картину, скульптуру и пр.), но в этом случае саккады совершаются в разных направлениях (вверх, вниз, в стороны и под углом) последовательно от одной точки объекта к другой.
Классическое изображение, описывающее саккадические движение глазных яблок
при рассматривании объекта
Система плавных (следящих) движений глазных яблок
Когда объект рассматривания перемещается, саккадическая система может первоначально зафиксировать его, но скоро теряет, поскольку изображение ускользает из области желтого пятна (сетчатое скольжение). Плавные (следящие) движения глаз необходимы для длительной фиксации движущегося объекта и слежения за ним. После того как визуальная цель выбрана, система работает вне волевого контроля.
Схематическое изображение функционирования системы
плавных (следящих) движений глаз
Вестибуло-окулярная система
В то время как система следящих движений глазных яблок фиксирует изображение перемещающегося объекта рассматривания на желтом пятне, существует другая система, которая позволяет стабилизировать изображение неподвижного объекта рассматривания на сетчатке во время движения головы. Это основная функция вестибуло-окулярной системы. Благодаря ее наличию у человека во время движения на транспорте по неровной дороге или ходьбе не возникает проблем с четким рассматриванием отдаленного объекта. В том случае, когда по какой-либо причине вестибуло-окулярная система не работает возникает феномен, называющийся “осциллопсия” – “дергание” визуальной картинки при движении.
Мозжечок
Основная функция мозжечка заключается в получении информации о положении тела в пространстве от всех органов чувств и регуляции на ее основе мышечного тонуса и движений для поддержания равновесия и выполнения точных действий.
Для больных с повреждением мозжечка характерна астазия-абазия – нарушение способности к сохранению равновесия тела при стоянии и ходьбе. Больные ходят, широко расставив ноги – так называемая туловищная атаксия (“пьяная походка”).
Ходьба на пятках и носках невозможна. Атаксия в данном случае развивается вследствие неспособности головного мозга координировать деятельность мышц в процессе преодоления силы тяжести. Также выявляются глазодвигательные расстройства. Они проявляются нарушением фиксации взора на неподвижных или двигающихся объектах, в результате чего возникают рывковые движения глаз при слежении. Также характерен вертикальный нистагм, бьющий вверх или вниз.
Вертикальный нистагм при повреждении мозжечка.
Средний мозг тонус мышц

Мышечный тонус определяется двумя факторами: механико-эластическими характеристиками мышечной и соединительной ткани; рефлекторной сократимостью мышц (тоническим рефлексом на растяжение). Первый фактор, или «внутренняя жесткость» мышечной ткани, имеет несомненное значение в развитии или поддержании спастического мышечного гипертонуса, поскольку в какой-то степени мышца подобна обыкновенной пружине, возвратная сила, действие которой пропорционально изменению ее длины и в то же время зависит от модуля упругости материала, из которого сделана пружина. Однако основная роль в поддержании и изменении мышечного тонуса отводится функциональному состоянию сегментарной дуги рефлекса растяжения (миотатического, или проприоцептивного рефлекса).
Рецепторным элементом миотатического рефлекса является инкапсулированное мышечное веретено. Каждая мышца содержит большое количество этих рецепторов. Мышечное веретено состоит из интрафузальных мышечных волокон (в сравнении с обычными экстрафузальными мышечными волокнами они значительно тоньше) и ядерной сумки, оплетенной спиралевидной сетью тонких нервных волокон, представляющих собой первичные чувствительные окончания. На некоторых интрафузальных волокнах имеются еще и вторичные, гроздевидные чувствительные окончания. При растяжении интрафузальных волокон первичные чувствительные окончания усиливают исходящую из них импульсацию, которая через быстро проводящие афферентные волокна типа Iа проводятся к альфа-большим мотонейронам спинного мозга. Оттуда через также быстропроводящие альфа-1-эфферентные волокна импульс идет к экстрафузальным белым мышечным волокнам, которые обеспечивают быстрое (фазическое) сокращение мышцы. От вторичных чувствительных окончаний, реагирующих на тонус мышцы, афферентная импульсация проводится по тонким волокнам II типа через систему вставочных нейронов к альфа-малым мотонейронам, которые иннервируют тонические экстрафузальные мышечные волокна (красные мышечные волокна), обеспечивающие поддержание тонуса и позы.
Гамма-мотонейроны находятся под влиянием центральных (супрасегментарных) воздействий, передающихся по волокнам, которые идут от мотонейронов оральных отделов головного мозга в составе пирамидного, ретикулоспинального, вестибулоспинального трактов. Таким образом, мышечный тонус может регулироваться непосредственно головным мозгом, что очень важно для выполнения произвольных движений. При этом если роль пирамидной системы заключается в преимущественной регуляции фазических (т.е. быстрых, целенаправленных) компонентов произвольных движений, то экстрапирамидная система обеспечивает плавность произвольных движений, их «настройку» в соответствии с решаемой задачей, т.е. преимущественно регулирует тоническую иннервацию мышечного аппарата.
В регуляции мышечного тонуса принимают участие и тормозные механизмы, которые реализуются с помощью рецепторов Гольджи, расположенных в сухожилиях мыщц, и вставочных клеток Реншоу, находящихся в передних рогах спинного мозга. Сухожильные рецепторы Гольджи при растяжении или значительном напряжении мышцы посылают афферентные импульсы, которые проводятся по быстропроводящим волокнам 1b типа в спинной мозг и оказывают тормозящее воздействие на мотонейроны передних рогов. Вставочные нейроны Реншоу активизируются через коллатерали при возбуждении альфа-мотонейронов и действуют по принципу отрицательной обратной связи, тормозя активность последних. Таким образом, нейрогенные механизмы регуляции мышечного тонуса многообразны и сложны.



