Скопление аксонов нейронов в спинном мозге
Взаиморасположение серого и белого вещества на разных уровнях спинного мозга представлено на рисунке ниже. Белое вещество представлено преимущественно аксонами и дендритами и образует задние, передние и боковые канатики (от лат. funiculus — «канатик») спинного мозга, которые затем разделяются на пучки нервных волокон (от лат. fascicule—«пучок»). Шейное (сегменты С5-Т1) и пояснично-крестцовое (сегменты L1-S2) утолщения образованы за счет увеличения объема серого вещества в данных сегментах, что необходимо для иннервации конечностей на противоположной стороне тела. Белое вещество больше представлено в верхних отделах спинного мозга, поскольку там проходят чувствительные и двигательные волокна, иннервирующие конечности.
Так, в заднем канатике спинного мозга располагается тонкий пучок, по которому передаются импульсы от нижних конечностей. Этот пучок представлен как в шейных, так и в пояснично-крестцовых сегментах. Напротив, клиновидный пучок, передающий импульсы от верхних конечностей, не представлен в поясничных сегментах спинного мозга.
Несмотря на то, что деление спинного мозга на пронумерованные сегменты (как указано выше) соответственно парам спинномозговых корешков довольно удобно для практического использования, фактически спинной мозг не имеет четкого сегментарного строения. Группы ядер, видимые на поперечных срезах спинного мозга, на самом деле представляют собой часть непрерывных клеточных столбиков, которые в большинстве случаев распространяются на несколько сегментов.
 Поперечные срезы спинного мозга на различных уровнях.
Поперечные срезы спинного мозга на различных уровнях.
а) Типы нейронов спинного мозга. Нейроны спинного мозга наименьшего размера (диаметром 5-20 нм) — промежуточные, их тела располагаются в спинном мозге. В то время как отростки одних промежуточных нейронов располагаются в пределах одного сегмента спинного мозга, аксоны других промежуточных нейронов распространяются в составе белого вещества спинного мозга на несколько сегментов вверх или вниз, тем самым обеспечивая связь между ними. Такие аксоны называют проприоспинальными (собственными) волокнами, которые формируют собственные пучки.
Большинство таких промежуточных нейронов участвует в формировании спинномозговых рефлексов. Другие промежуточные нейроны располагаются между волокнами нисходящих путей и мотонейронами, участвующими в регуляции двигательной активности. Кроме того, функция некоторых промежуточных нейронов заключается в проведении чувствительных импульсов от низших уровней ЦНС к высшим.
Наиболее часто встречаемый тип нейронов в сером веществе — нейроны среднего размера (диаметром 20-50 нм). Большинство из них выполняет функцию промежуточного звена (ретрансляции) при передаче афферентных импульсов от задних корешков к головному мозгу посредством аксонов, которые образуют тракты. Трактом (проводящим путем) считают совокупность нервных волокон, выполняющих единую функцию. Как будет показано далее, термин «тракт» часто употребляют некорректно, поскольку изначально полагают, что входящие в состав группы волокна выполняют одну функцию, однако на самом деле данная группа представлена волокнами разных функциональных классов.
Наиболее крупные нейроны в спинном мозге — а-мотонейроны (диаметром 5-20 нм), которые иннервируют скелетные мышцы. Среди них в сером веществе передних рогов диффузно располагаются меньшие по размеру у-мотонейроны, осуществляющие эфферентную иннервацию нервно-мышечных веретен. В средней части передних рогов находятся клетки Реншоу, функция которых заключается в торможении а-мотонейронов.
Спинномозговые рефлекторные дуги, которые начинаются от мышечных веретен и рецепторов сухожилий, защитный рефлекс описна в отдельных статьях на сайте.
На основе цитоархитектонических характеристик (например, размер нейронов, особенности окраски, наличие рецепторов и нейронных связей) серое вещество спинного мозга принято разделять на 10 слоев (спинномозговых пластинок), называемых пластинами Рекседа. Эти пластины были выделены с описательной целью, и не всегда какой-либо пластине соответствует определенная функция. Структура пластин меняется в зависимости от изучаемого уровня спинного мозга: в то время как на одном уровне в пределах пластины можно наблюдать определенные ядра, на другом уровне они будут выражены не так явно.
 Два сегмента спинного мозга. Изображена столбчатая организация ядер переднего рога спинного мозга.
Два сегмента спинного мозга. Изображена столбчатая организация ядер переднего рога спинного мозга. 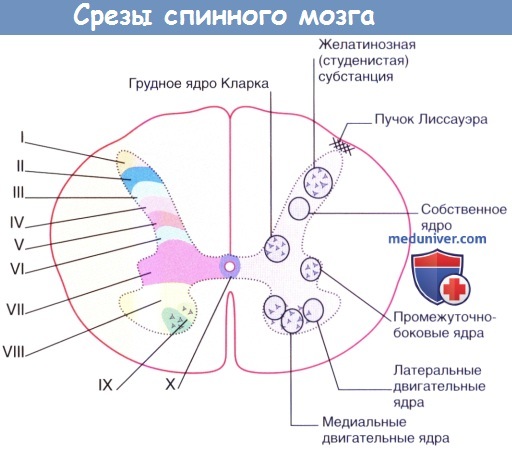 Спинномозговые пластинки (пластины Рекседа) (I-X) и скопления тел нейронов (ядра) на среднем грудном уровне спинного мозга.
Спинномозговые пластинки (пластины Рекседа) (I-X) и скопления тел нейронов (ядра) на среднем грудном уровне спинного мозга.
б) Спинальные ганглии. Спинальные, или заднекорешковые, ганглии (спинномозговые узлы) расположены по ходу задних корешков спинного мозга в области межпозвоночных отверстий. В этой области передние и задние корешки соединяются и формируют спинномозговые нервы. Грудные спинальные ганглии содержат около 50000 униполярных нейронов, а проводящие пути чувствительной иннервации от верхних и нижних конечностей — около 100000. От тела униполярных (или, точнее, псевдоуниполярных) нейронов отходит только один отросток — короткий стволовой аксон. В связи с этим аксоны и дендриты этих нейронов морфологически неотличимы. Отдельные клетки ганглия окружены модифицированными шванновскими клетками — амфицитами (клетками-сателлитами,или мантийными глиоцитами).
1. Центральные окончания чувствительных нервных волокон. В зоне вступления волокон заднего корешка в спинной мозг чувствительные нервные волокна делятся на внутренние и наружные пучки. Внутренний пучок содержит крупные и средние волокна, которые в дальнейшем в пределах заднего канатика делятся на восходящие и нисходящие ветви. Затем ход волокон ветвей отклоняется в сторону заднего рога спинного мозга, где часть из них образует синапсы в области заднего ядра (ядра Кларка). Наиболее крупные восходящие волокна поднимаются до ядер задних столбов (тонкое/клиновидное) в продолговатом мозге, формируя при этом основную часть нервных волокон в составе тонкого и клиновидного пучков.
Наружный пучок образован небольшими волокнами (А δ-и С-волокна), которые при вступлении в спинной мозг делятся на восходящие и нисходящие ветви в области пучка Лиссауэра и образуют синапсы с нейронами в составе желатинозной субстанции. Некоторые волокна образуют синапсы с дендритами нейронов в собственном ядре, с которого начинается спинно-таламический проводящий путь.
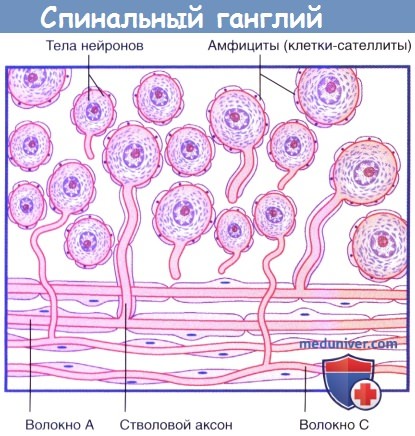 Спинальный ганглий.
Спинальный ганглий.
Обратите внимание на Т-образное разделение волокон стволового аксона в нижней части рисунка:
именно по этой причине эти нейроны называют «псевдоу-ниполярными». 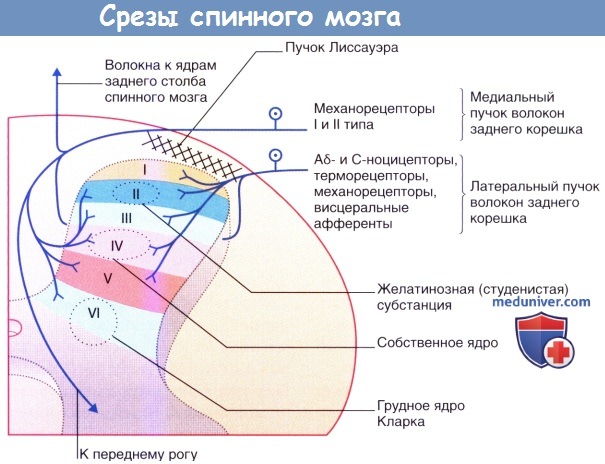 Окончания первичных чувствительных нейронов на уровне заднего рога спинного мозга.
Окончания первичных чувствительных нейронов на уровне заднего рога спинного мозга.
Редактор: Искандер Милевски. Дата публикации: 15.11.2018
Скопление аксонов нейронов в спинном мозге
· Двигательные функции спинного мозга
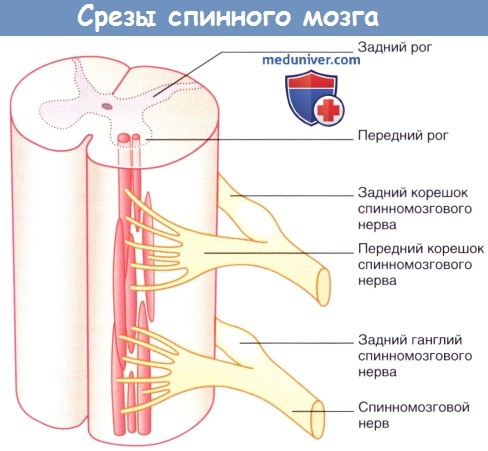
Спинной мозг (см. рис. 9–2) покрыт мягкой, паутинной и твёрдой мозговой оболочкой, омывается спинномозговой жидкостью и состоит из двух симметричных половин, соединённых узкой перемычкой, содержащей центральный канал. На поперечном разрезе легко различимы лежащее снаружи белое вещество и расположенная глубже тёмная масса — серое вещество. В спинной мозг входят проходящие в составе задних корешков центральные отростки чувствительных нейронов спинномозговых узлов. В передних рогах находятся мотонейроны, аксоны которых через передние корешки иннервируют скелетные мышцы. Передний и задний корешки соединяются и образуют 31 пару спинномозговых нервов.
Ядра и проводящие пути спинного мозга
Карта расположения путей и ядер спинного мозга показана на рис. 13–1.
Серое вещество на поперечном разрезе имеет характерную форму бабочки, состоит из отростков нервных клеток и их перикарионов, образующих скопления — ядра, объединённые в пластинки. Каждая половина серого вещества формирует на протяжении всего спинного мозга выступы — серые столбы: передний столб — columna anterior, задний столб — columna posterior и боковой столб — columna lateralis. Столб на поперечном разрезе получает название рога, соответственно передний (cornu anterius), задний (cornu posterius) и боковой (cornu laterale). Между передним и задним рогами в составе серого вещества различают промежуточную зону (substantia intermedia, пластинка VII). Правая и левая половины серого вещества соединены серой спайкой (comissura grisea), находящейся в пределах пластинки X. Центральный канал (canalis centralis) разделяет серую спайку на переднюю (comissura grisea anterior) и заднюю (comissura grisea posterior) части.
· Пластинки (рис. 1 3 –1, табл. 1 3 –1). Перикарионы нейронов серого вещества по длине спинного мозга картированы по десяти пластинкам. Топография ядер соответствует топографии пластинок, хотя они не всегда совпадают.
à Пластинка I. Тонкий слой серого вещества у поверхности заднего столба; содержит мелкие нейроны, нейроны средних размеров, а также крупные веретеновидные нейроны, расположенные параллельно поверхности серого вещества. Здесь, в пределах N. dorsomarginalis, найдены безмиелиновые аксоны, короткие дендриты и синаптические контакты. Перикарионы нейронов образуют синаптические контакты с аксонами из пластинки II, а аксоны псевдоуниполярных нейронов оканчиваются на дендритах части нейронов пластинки I. Основная часть аксонов, поступающих из пластинки II, а также принадлежащих псевдоуниполярным нейронам, входит в состав краевого пояса Лиссауэра (fasciculus dorsolateralis). Нейроны пластинки I реагируют на болевые и температурные стимулы и отдают волокна спиноталамическому пути противоположной стороны. В пластинке I присутствуют нейроны, содержащие вещество Р и энкефалин, и проходят волокна, дающие положительную иммуноцитохимическую реакцию на вещество Р, энкефалин, соматостатин и серотонин.
à Пластинка II располагается вентро-медиально от пластинки I, на всех уровнях спинного мозга содержит скопление мелких нейронов — роландово студенистое вещество (substantia gelatinosa Rolandi). В пластинку II входят аксоны из краевого пояса Лиссауэра, заднего канатика и прилегающих частей бокового канатика. К нейронам внутренней зоны пластинки II подходят волокна болевой и температурной чувствительности, а к нейронам наружной зоны — волокна тактильной чувствительности. Аксоны нейронов пластинки II уходят в краевой пояс Лиссауэра и собственные пучки. Другая часть аксонов заканчивается в пределах той же пластинки на других уровнях. Нейроны пластинки II влияют на возбудимость крупных нейронов центральных отделов серого вещества (например, пластинки IV), дендриты которых проникают в пластинку II и контактируют с перикарионами её нейронов. Пластинка II, как и пластинка I, содержит в большом количестве вещество Р — нейромедиатор передачи возбуждения от терминалей центральных отростков псевдоуниполярных нейронов спинномозговых узлов к вставочным нейронам спинного мозга, а также энкефалин.
à Пластинка III. Ветви дендритов нервных клеток этой пластинки оканчиваются в пластинках II и I; аксоны дихотомически делятся и образуют густое сплетение в пластинках III и IV.
à Пластинка IV. Нейроны отвечают на тактильные сигналы, возникающие при лёгком прикосновении. Аксоны некоторых нейронов в составе проекционных путей переходят через переднюю белую спайку на противоположную сторону и восходят к таламусу.
à Пластинка V расположена в основании заднего рога. Латеральная часть пластинки участвует в образовании ретикулярной формации, наиболее выраженной в шейном отделе. Нейроны варьируют по размерам и форме. Дендриты некоторых из них направляются в пластинку II, где вступают в контакт с центральными отростками псевдоуниполярных нейронов.
à Пластинка VI присутствует только в утолщениях спинного мозга (intumescentia cervicalis et lumbalis). В медиальной части пластинки оканчиваются мышечные афференты, а в латеральной части проходят нисходящие спинальные пути.
à Пластинка VII (zona intermedia) расположена между передними и задними рогами. В утолщениях пластинка заходит в область передних рогов. Нейроны пластинки образуют несколько ядер. Нейроны, не входящие в состав ядер данной пластинки, вместе с нейронами прилежащих областей пластинок V и VI образуют перекрещивающиеся волокна переднего спиномозжечкового пути. Здесь различают несколько ядер (табл. 13–1).
Скопление аксонов нейронов в спинном мозге




§ 30. Спинной мозг
Спинной мозг человека состоит из 31 сегмента.
В первые три месяца зародышевой жизни спинной мозг располагается на протяжении всего позвоночного канала, но позднее занимает только часть его длины, так как растет медленнее позвоночного столба. Поэтому корешки спинномозговых нервов, появляющиеся из мозга вначале на уровне межпозвоночных отверстий соответствующего сегмента, оказываются впоследствии выше своих отверстий и по выходе из мозга тянутся на некотором протяжении внутри позвоночного канала. Особенно большое расстояние проходят корешки поясничного и крестцового отделов спинного мозга, благодаря чему из них образуется так называемый конский хвост.
Серое вещество. На разрезе спинного мозга видно, что его очень узкий центральный канал окружен серым веществом, выступающие части которого образуют передние и задние рога (рис. 115). В грудном отделе и в верхней части поясничного между передним и задним выдаются еще и боковые рога. На периферии расположено белое вещество. В последнем между задним и передним (или боковым) рогами находятся переплетающиеся перекладины серого вещества, выделяемые в качестве сетчатого вещества, или ретикулярной формации (рис. 115).
По срединной линии передней поверхности мозга тянется передняя срединная щель, разделяющая его на правую и левую половины. Борозда не достигает серого вещества; между ним и ее дном находится белая спайка, соединяющая половины мозга.
Стенки задней срединной борозды сращены, и на ее месте через всю толщу белого вещества проходит глиальная перегородка.
В непосредственной близости от серого вещества, внутри всех столбов, лежат короткие межсегментальные волокна основных пучков (Атл., 73, внизу). Начавшись от клеток ретикулярной формации, волокна проходят вверх и вниз 2-3 сегмента и оканчиваются на мотонейронах передних рогов. По волокнам устанавливается связь между отдельными сегментами, поэтому основные пучки выделяются в собственный аппарат спинного мозга.
Волокна спинномозговых ганглиев, проникающие в мозг в составе дорсальных корешков, продолжают свой путь по различным направлениям. Одни из волокон оканчиваются на мотонейронах переднего рога своего сегмента, на вставочных нейронах задних рогов своей или противоположной стороны, на нейронах автономной нервной системы боковых рогов и на клетках ретикулярной формации. В результате в спинном мозге осуществляются простейшие (безусловные) рефлексы со всех сегментов тела в ответ на раздражения кожи, мышц и внутренних органов. Другие волокна поднимаются вверх, входя в состав задних канатиков; они относятся к восходящим проводящим путям спинного мозга.
К восходящим путям спинного мозга относятся тонкий и клиновидный пучки, задний и передний спинно-мозжечковые пути, боковой спинно-таламический и др. (Атл., 73, внизу).
Тонкий и клиновидный пучки проходят в задних канатиках и образованы нейритами чувствительных нейронов спинальных ганглиев.
Задний спинно-мозжечковый путь лежит в боковых канатиках. Он берет начало от клеток ядра, которое находится в основании задних рогов (дорсальное ядро) одноименной стороны.
Передний спинно-мозжечковый путь состоит из отростков вставочных нейронов задних рогов. После перекреста средней линии мозга волокна входят в состав боковых канатиков противоположной стороны.
Оба пути проводят проприорецепторные импульсы к мозжечку.
Боковой спинно-бугорный путь находится также в боковых канатиках и состоит из перекрещенных волокон вставочных нейронов заднего рога противоположной стороны. Путь проводит импульсы болевой и температурной чувствительности тела к межуточному мозгу.
Перекрест восходящих проводящих путей, совершаемый обычно волокнами вставочных нейронов, приводит к тому, что импульс попадает в полушарие, противоположное той стороне тела, от которой идет возбуждение.
Нисходящие пути состоят из красноядерно-спинального, бокового и переднего корково-спинномозговых, текто-спинномозгового, преддверно-спинномозгового, медиального продольного пучка и др. (Атл., 73, внизу).
Красноядерно-спинномозговой путь начинается из среднего мозга (от красного ядра), спускается по боковому канатику противоположной стороны спинного мозга и оканчивается на двигательных нейронах передних рогов. Несет непроизвольные двигательные импульсы.
Боковой корково-спинномозговой путь лежит в боковом канатике и состоит из нейритов клеток коры противоположного полушария. Путь постепенно истончается, так как в каждом сегменте спинного мозга часть его волокон заканчивается на клетках передних рогов. Путь проводит от коры произвольные двигательные импульсы, стимулирующие и тормозные.
Передний корково-спинномозговой путь, как и боковой, состоит из волокон клеток коры больших полушарий, но лежит в переднем канатике. Волокна его оканчиваются на мотонейронах главным образом противоположной стороны, переходя туда в составе передней спайки спинного мозга. Этот путь имеет ту же функцию, что и боковой корково-спинномозговой.
Представляет интерес, что корково-спинальные пути оканчиваются на мотонейронах спинного мозга лишь у человека и приматов, в то время как у субприматов, а иногда и у приматов между ними включается вставочный нейрон (Атл., 73, внизу слева). Функциональное обоснование этому явлению не найдено.
Текто-спинномозговой путь лежит также в переднем канатике, начинается, от верхних и нижних холмиков крыши среднего мозга и оканчивается на клетках передних рогов.
Преддверно-спинномозговой путь лежит между передним и боковым канатиками. Он идет от продолговатого мозга к передним рогам и проводит импульсы, обеспечивающие равновесие тела.
Медиальный продольный пучок лежит в переднем канатике и состоит как из нисходящих, так и восходящих волокон; берет начало и оканчивается на ядрах ствола мозга и на клетках передних рогов. Пучок представляет собой очень древнюю систему волокон, которая у низших позвоночных служит важнейшим ассоциационным путем головного мозга.
Большинство нисходящих и восходящих путей совершает перекрест на разных уровнях центральной нервной системы. В результате импульс, пройдя на протяжении всей рефлекторной дуги два перекреста (в восходящем и нисходящем направлениях), возвращается на сторону, получившую раздражение.
Сосуды спинного мозга. Спинной мозг питается из сегментальных межреберных, поясничных и подключичных артерий, главным образом через позвоночные артерии. Войдя в полость черепа через большое затылочное отверстие, позвоночные артерии отдают две парные передние и задние спинномозговые артерии (Атл., 74). Передние вскоре соединяются в непарный стволик, идущий вниз вдоль передней срединной щели; задние спускаются по линии вхождения в спинной мозг его задних корешков. Все три артерии связаны друг с другом многочисленными анастомозами и получают подкрепление из указанных выше сегментальных сосудов. В глубь мозга артерии посылают тонкие веточки; серое вещество снабжается кровью лучше, чем белое. Кровоотток из спинного мозга осуществляется по венам, сопровождающим артерии и впадающим в венозное сплетение эпидурального пространства.
Скопление аксонов нейронов в спинном мозге
Центральная нервная система
Спинной мозг
Развитие спинного мозга. Как уже говорилось выше, филогенетически спинной мозг (туловищный мозг ланцетника) появляется на III этапе развития нервной системы (трубчатой нервной системы). В это время головного мозга еще нет, поэтому туловищный мозг имеет центры для управления всеми процессами организма, как вегетативными, так и анимальными (висцеральные и соматические центры). Соответственно сегментарному строению тела туловищный мозг имеет сегментарное строение, он состоит из связанных между собой невромеров, в пределах которых замыкается простейшая рефлекторная дуга. Метамерное строение спинного мозга сохраняется и у человека, чем и обусловливается наличие у него коротких рефлекторных дуг.
С появлением головного мозга (этап цефализации) в нем возникают высшие центры управления всем организмом, а спинной мозг попадает в подчиненное положение. Спинной мозг не остается только сегментарным аппаратом, а становится и проводником импульсов от периферии к головному мозгу и обратно, и в нем развиваются двусторонние связи с головным мозгом. Таким образом, в процессе эволюции спинного мозга образуются два аппарата: один, более старый, сегментарный аппарат собственных связей спинного мозга и второй, более новый, надсегментарный аппарат двусторонних проводящих путей к головному мозгу. Такой принцип строения наблюдается и у человека.
Решающим фактором образования туловищного мозга является приспособление к окружающей среде при помощи движения. Поэтому строение спинного мозга отражает способ передвижения животного. Так, например, у пресмыкающихся, не имеющих конечностей и передвигающихся с помощью туловища (например, у змеи), спинной мозг развит равномерно на всем протяжении и не имеет утолщений. У животных, пользующихся конечностями, возникает два утолщения; при этом, если более развиты передние конечности (например, крылья птиц), то преобладает переднее (шейное) утолщение спинного мозга; если более развиты задние конечности (например, ноги страуса), то увеличено заднее (поясничное) утолщение; если в ходьбе одинаково участвуют и передние, и задние конечности (четвероногие млекопитающие), то одинаково развиты оба утолщения. У человека в связи с более сложной деятельностью руки как органа труда шейное утолщение спинного мозга дифференцировалось сильнее, нежели поясничное.
Отмеченные факторы филогенеза играют роль в развитии спинного мозга и в онтогенезе. Спинной мозг развивается из мозговой трубки, из ее заднего отрезка (из переднего возникает головной мозг.). Из вентрального отдела мозговой трубки образуются передние столбы серого вещества спинного мозга (клеточные тела двигательных нейронов), прилегающие к ним пучки продольных нервных волокон и отростки названных нейронов (двигательные корешки). Из дорсального отдела возникают задние столбы серого вещества (клеточные тела чувствительных нейронов), задние канатики (отростки чувствительных нейронов).
Из-за редукции каудальной части спинного мозга получается тонкий тяж из нервной ткани, будущая Шит terminate. Первоначально, на 3-м месяце утробной жизни, спинной мозг занимает весь позвоночный канал, затем позвоночник начинает расти скорее, чем мозг, вследствие чего конец последнего постепенно перемещается кверху (краниально). При рождении конец спинного мозга уже находится на уровне III поясничного позвонка, а у взрослого достигает высоты I-II поясничного позвонка. Благодаря такому «восхождению» спинного мозга отходящие от него нервные корешки принимают косое направление (рис. 267).
Строение спинного мозга
Спинной мозг, medulla spinalis (греч. myelos) лежит в позвоночном канале и у взрослых представляет собой длинный (45 см у мужчин и 41-42 см у женщин), несколько сплюснутый спереди назад цилиндрический тяж, который вверху (краниально) непосредственно переходит в продолговатый мозг, а внизу (каудально) оканчивается коническим заострением, conus medullaris, на уровне II поясничного позвонка (см. рис. 267). Знание этого факта имеет практическое значение (чтобы не повредить спинной мозг при поясничном проколе с целью взятия спинномозговой жидкости или с целью спинномозговой анестезии, надо вводить иглу шприца между остистыми отростками III и IV поясничных позвонков).
От conus medullaris отходит книзу так называемая концевая нить, filum terminate, представляющая атрофированную нижнюю часть спинного мозга, которая внизу состоит из продолжения оболочек спинного мозга и прикрепляется ко II копчиковому позвонку.
* ( По последним данным (Гассер, 1955; Гесс, 1956; Митчелл, 1957, и др.), в задних корешках проходят безмякотные эфферентные волокна, иннервирующие гладкие мышцы внутренностей и сосудов.)
* ( В составе спинальных ганглиев могут встретиться мультиполярные элементы вегетативной нервной системы.)
Вследствие того, что спинной мозг короче позвоночного канала, место выхода нервных корешков не соответствует уровню межпозвонковых отверстий. Чтобы попасть в последние, корешки направляются не только в стороны от мозга, но еще и вниз и при этом тем отвеснее, чем ниже они отходят от спинного мозга. В поясничной части последнего нервные корешки спускаются к соответствующим межпозвонковым отверстиям параллельно filum terminate, облекая ее conus medullaris густым пучком, который носит название конского хвоста, cauda equina (см. рис. 267).
Внутреннее строение спинного мозга. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого вещества, слагающегося из миелиновых нервных волокон.
Серое вещество, окружающее центральный канал, носит название промежуточного вещества, substantia intermedia centralis. В каждой колонне серого вещества различают два столба: передний, columna grisea anterior, и задний, columna grisea posterior.
На поперечных разрезах спинного мозга эти столбы имеют вид рогов: переднего, расширенного, cornu anterius, и заднего, заостренного, cornu posterius. Благодаря этому общий вид серого вещества, выделяющегося на фоне белого, напоминает букву «Н».
Заложенные в заднем роге клетки образуют вторые, вставочные, нейроны; они дают начало нейритам, идущим в головной мозг, а клетки студенистого вещества и диффузно разбросанные в сером веществе рассеянные клетки, так называемые пучковые клетки, служат для связи с третьими нейронами, заложенными в передних рогах того же сегмента. Отростки этих клеток, идущие от задних рогов к передним, естественно, располагаются вблизи серого вещества, по его периферии, образуя узкую кайму белого вещества, непосредственно окружающего серое со всех сторон. Это собственные, или основные, пучки спинного мозга, fasciculi proprii. Аксоны других пучковых клеток делятся на восходящую и нисходящую ветви, которые оканчиваются на клетках передних рогов нескольких выше и нижележащих сегментов. Вследствие этого раздражение, идущее из определенной области тела, может передаваться не только на соответствующий ей сегмент спинного мозга, но захватывать и другие. В результате простой рефлекс может вовлекать в ответную реакцию целую группу мышц, обеспечивая сложное координированное движение, остающееся, однако, безусловно-рефлекторным.
* ( Клетки передних рогов являются также и трофическими центрами мускулатуры; выключение двигательных нейронов влечет за собой не только паралич, но и атрофию мышц.)
Наибольшее число ядер содержится в передних рогах шейного утолщения спинного мозга, откуда иннервируются верхние конечности, что определяется участием последних в трудовой деятельности человека. У последнего в связи с усложнением движений руки как органа труда этих ядер значительно больше, чем у животных, включая антропоидов. Таким образом, задние и передние рога серого вещества имеют отношение к иннервации органов животной жизни, особенно аппарата движения, в связи с усовершенствованием которого в процессе эволюции и развивался спинной мозг.
Передний и задний рога в каждой половине спинного мозга связаны между собой промежуточной зоной серого вещества, которая в грудном и поясничном отделах спинного мозга, на протяжении от I грудного до II-III поясничных сегментов, особенно выражена и выступает в виде бокового рога, cornu laterale. Вследствие этого в названных отделах серое вещество на поперечном разрезе приобретает вид бабочки. В боковых рогах заложены клетки, иннервирующие вегетативные органы и группирующиеся в ядро, которое носит название nucleus intermediolateralis (впервые описано И. М. Якубовичем). Нейриты клеток этого ядра выходят из спинного мозга в составе передних корешков.
Б. Белое вещество, substantia alba, спинного мозга состоит из нервных отростков, которые составляют три системы нервных волокон:
1. Короткие пучки ассоциационных волокон, соединяющих участки спинного мозга на различных уровнях (афферентные и вставочные нейроны).
2. Длинные центростремительные (чувствительные, афферентные) волокна.
3. Длинные центробежные (двигательные, эфферентные) волокна.
Первая система (коротких волокон) относится к собственному аппарату спинного мозга, а остальные две системы (длинных волокон) составляют проводниковый аппарат двусторонних связей с головным мозгом.
Аппарат двусторонних связей с головным мозгом является филогенетически более молодым, так как он возник лишь тогда, когда появился головной мозг.
По мере развития последнего разрастались кнаружи и проводящие пути, связывающие спинной мозг с головным (рис. 269). Этим объясняется тот факт, что белое вещество спинного мозга как бы окружило со всех сторон серое вещество. Благодаря проводниковому аппарату собственный аппарат спинного мозга связан с аппаратом головного мозга, который объединяет работу всей нервной системы. Нервные волокна группируются в пучки, различимые на препарате лишь с помощью особых методов (см. курс микроскопической анатомии), а из пучков составляются видимые невооруженным глазом канатики: задний, боковой и передний. В заднем канатике (рис. 270), прилежащем к заднему (чувствительному) рогу, лежат пучки восходящих нервных волокон; в переднем канатике, прилежащем к переднему (двигательному) рогу, лежат пучки нисходящих нервных волокон; наконец, в боковом канатике находятся и те и другие. Кроме канатиков, белое вещество находится в белой спайке, comissura alba, образующейся вследствие перекреста волокон спереди от substantia intermedia centralis; сзади белая спайка отсутствует.
Задние канатики содержат волокна задних спинномозговых корешков, слагающиеся в две системы:
1. Медиально расположенный нежный пучок Голля, fasciculus gracilis (Goll).
2. Латерально расположенный клиновидный пучок Бурдаха fasciculus cunedtus (Burdach).
Боковые канатики содержат следующие пучки:
К заднему мозгу: 1) tractus spinocerebellaris posterior (Flechsig), задний спинно-мозжечковый путь, располагается в задней части бокового канатика по его периферии; 2) tractus spinocerebellaris anterior (Gowers), передний спинно-мозжечковый путь, лежит вентральнее предыдущего.
Оба спинно-мозжечковых тракта проводят бессознательные проприоцептивные импульсы (бессознательная координация движений).
К среднему мозгу: 3) tractus spinotectalis прилегает к медиальной стороне и передней части tractus spinocerebellaris anterior.
От коры большого мозга: 1) боковой пирамидный путь, tr. corticospinal (pyramidalis) lateralis. Этот тракт является сознательным эфферентным двигательным путем.
От среднего мозга: 2) tractus rubrospindlis (Monakow). Он является бессознательным эфферентным двигательным путем.
Передние канатики содержат нисходящие пути.
От коры головного мозга: 1) передний пирамидный тракт, iractus corticospinal (pyramidalis) anterior. Путь составляет с боковым пирамидным пучком общую пирамидную систему.
Ряд пучков идет к передним рогам спинного мозга от различных ядер продолговатого мозга, имеющих отношение к равновесию и координации движений, а именно:
5) собственные пучки, fasciculi proprii, непосредственно прилегают к серому веществу и относятся к собственному аппарату спинного мозга.
Оболочки спинного мозга
Спинной мозг одет тремя соединительнотканными оболочками, meninges, происходящими из мезодермы вокруг мозговой трубки. Оболочки эти следующие, если идти с поверхности вглубь: твердая оболочка, dura mater, или pachymeninx; паутинная оболочка, arachnoidea, и сосудистая оболочка, pia mater. Последние две оболочки в противоположность первой называются еще мягкой оболочкой, leptomeninx. Краниально все три оболочки продолжаются в такие же оболочки головного мозга.
Артерии свои твердая оболочка получает от спинномозговых ветвей сегментарных артерий, вены ее вливаются в plexus venosus vertebralis internus, а нервы ее происходят от rami meningei спинномозговых нервов. Внутренняя поверхность твердой оболочки покрыта слоем эндотелия, вследствие чего имеет гладкий, блестящий вид.
2. Паутинная оболочка спинного мозга, arachnoidea spinalis, в виде тонкого прозрачного бессосудистого листка прилегает изнутри к твердой оболочке, отделяясь от последней щелевидным, пронизанным тонкими перекладинками, субдуральным пространством, cavum subdural. Между паутинной оболочкой и непосредственно покрывающей спинной мозг сосудистой оболочкой находится подпаутинное пространство, cavum subarachnoideale, в котором мозг и нервные корешки лежат свободно, окруженные большим количеством спинномозговой жидкости, liquor cerebrospinal. Это пространство в особенности широко в нижней части арахноидального мешка, где оно окружает cauda equina спинного мозга (cisterna terminalis). Наполняющая подпаутинное пространство жидкость находится в непрерывном сообщении с жидкостью подпаутинных пространств головного мозга и мозговых желудочков. Между паутинной оболочкой и покрывающей спинной мозг сосудистой оболочкой в шейной области сзади вдоль средней линии образуется перегородка, septum cervicdle intermedium. Кроме того, по бокам спинного мозга во фронтальной плоскости располагается зубчатая связка, tig. denticuldtum, состоящая из 19-23 зубцов, проходящих*^ промежутках между передними и задними корешками. Зубчатые связки служат для укрепления мозга на месте, не позволяя ему вытягиваться в длину. Посредством обеих ligg. denticulata подпаутинное пространство делится на передний и задний отделы.
3. Сосудистая оболочка спинного мозга, pfa mater spinalis, покрытая с поверхности эндотелием, непосредственно облекает спинной мозг и содержит между 2 своими листками сосуды, вместе с которыми заходит в его борозды и мозговое вещество, образуя вокруг сосудов периваскулярные лимфатические пространства.
Сосуды спинного мозга (рис. 271). Аа. spinales anterior et posteriores, спускаясь вдоль спинного мозга, соединяются между собой многочисленными ветвями, образуя на поверхности мозга сосудистую сеть (так называемую vasocorona). От этой сети отходят веточки, проникающие вместе с отростками сосудистой оболочки в вещество мозга (см. рис. 271) (И. Д. Лев). Вены в общем аналогичны артериям и впадают в конечном итоге в plexus venosi vertebrates interni. К лимфатическим сосудам спинного мозга можно отнести периваскулярные пространства вокруг сосудов, сообщающиеся с подпаутинным пространством.
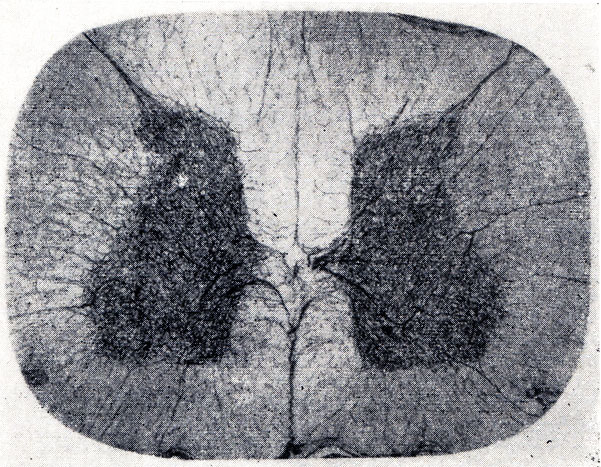
Рис. 271. Сосуды спинного мозга кошки (по И. Д. Лев)



