Численность и рост популяции: факторы регуляции численности популяции
Содержание:
Численность популяций любого вида в природе не остаётся неизменной. Колебания численности в первую очередь связаны с пищей, убежищами, местами для размножения. Неограниченному росту численности популяции препятствует нехватка ресурсов для существования на любой территории. Отклонения показателей численности от оптимального (среднего) уровня вызывают многие факторы окружающей среды. Возвращение к среднему показателю в биологии называется регуляцией.
Популяции

Для характеристики популяции используются такие показатели, как численность (общее число организмов данного вида в местообитании), рождаемость, смертность. Один из основных параметров — плотность или количество особей на единицу поверхности (в единице объёма).
Важны и другие показатели:
При изучении конкретной популяции обращают внимание на её численность и плотность. Пример: численность волков может быть ограничивающим фактором для зайцев. Волк питается травоядными животными, а при высокой численности хищников им требуется больше корма. Численность популяции зайцев может сильно сократиться. И наоборот, сокращение численности хищников ведёт к увеличению численности популяции жертв — размножению грызунов, распространению среди них инфекционных заболеваний.
При чрезмерном увеличении численности и плотности популяции возрастает сила давления, оказываемая на среду обитания и другие живые организмы. При сокращении численности существует опасность исчезновения данного вида, сокращения биоразнообразия. Кроме того, экологическую нишу исчезающего вида может занять более агрессивный.
Если условия оптимальные для популяции, то численность организмов данного вида увеличивается с максимальной скоростью. Так происходит при обилии пищи, отсутствии врагов и перенаселении самой популяции. Примеры таких изменений: «цветение» воды в стоячих водоёмах из-за усиленного размножения водорослей, вспышки размножения саранчи.
Неоднородность популяции — это фактор благоприятный для выживания. Наоборот, внутри- и межвидовая конкуренция в условиях нехватки ресурсов приводят к замедлению роста популяций. В какой-то момент времени численность достигает уровня, когда рождаемость равна смертности. Такое состояние называется равновесной плотностью популяции.
Как регулируется численность популяции
В природе никогда не происходит неограниченный рост численности. Действуют лимитирующие факторы, поэтому виды не размножаются бесконечно. Суммарные потребности особей одного вида в определённом ресурсе могут превышать его количество в экосистеме. Организмы начинают конкурировать за ресурс, а те, что его недополучают, слабеют, перестают размножаться либо гибнут.
Рассмотрим факторы, влияющие на численность популяций: не зависящие и зависящие от её плотности. Не зависящими от плотности считаются такие факторы, на действие которых не влияет изначальная численность и плотность популяции. К этой группе относятся сильные морозы, засухи, ураганы, наводнения. Число погибших и выживших особей в результате природной стихии не зависит от первоначальной численности и плотности популяции.
Разберём задание: «Какой абиотический фактор среды вызывает миграцию птиц?»
Перелёты пернатых связаны не с одним фактором, а с природным механизмом регуляции сезонных изменений в жизни птиц. Уменьшается количество корма, его становится недостаточно для всех особей, усиливаются внутри- и межвидовая конкуренции. Сигналом к началу миграции является изменение длины дня, что приводит к гормональным изменениям в организме перелётных птиц.
Факторы, зависящие от плотности, сильнее влияют на популяцию по мере роста её численности и увеличения плотности. К этой группе относятся биотические факторы — внутри- и межвидовые. Чем выше плотность особей, тем сильнее действуют эти факторы.
Зависят от плотности:
Действие этих факторов повышается по мере увеличения плотности, что препятствует в дальнейшем неблагоприятному росту численности популяции. Наблюдается отрицательная зависимость или обратная связь.
2. Демографические показатели популяции
Теория:
Важно понимать причины изменения популяций и скорость этих изменений, а также уметь определять их групповые характеристики, т. е. демографические показатели.
Рассмотрим эти показатели подробнее.
Плотность используется, если нужно знать динамику численности популяции, т.е. изменение количества особей во времени.
Рождаемость определяет возможность развития популяции за счёт размножения населяющих её особей.
Различают максимальную рождаемость (иногда её называют физиологической, или абсолютной) и экологическую, или просто рождаемость.
Максимальная ( потенциальная ) рождаемость — это наибольшее значение скорости появления новых особей, теоретически возможное при отсутствии факторов, ограничивающих этот процесс, т. е. в идеальных условиях.
Максимальная рождаемость определяется плодовитостью самок, т. е. их способностью производить определённое количество потомства, а также общим количеством потомков (за определённый срок).
Экологическая рождаемость — это число новых особей, появившихся в популяции за единицу времени; она позволяет судить о скорости возрастания численности популяции при данных условиях жизни рассматриваемой группы особей.
Экологическая рождаемость непостоянна и изменяется в зависимости от физических условий среды и состава популяции.
Смертностью определяется средняя продолжительность жизни, характеризующая группу особей. Средняя продолжительность жизни больше в той группе, где смертность ниже.
СОДЕРЖАНИЕ
Обзор: Типы эффективной численности населения
Эмпирические измерения
В популяциях дрозофилы численностью 16 по данным переписи эффективный размер дисперсии равен 11,5. Это измерение было достигнуто путем изучения изменений частоты нейтрального аллеля от одного поколения к другому в более чем 100 реплицированных популяциях.
Для получения слитых эффективных размеров популяций, обзор публикаций по 102 в основном диких животных видов животных и растений дали 192 Н е / N коэффициентов. В обследованных исследованиях использовалось семь различных методов оценки. Соответственно, это соотношение широко варьировалось от 10 -6 для тихоокеанских устриц до 0,994 для человека, в среднем 0,34 для исследованных видов. Генеалогический анализ людей-охотников-собирателей ( эскимосов ) определил соотношение эффективной численности популяции к переписной для гаплоидных (митохондриальная ДНК, Y-хромосомная ДНК) и диплоидных (аутосомная ДНК) локусов по отдельности: отношение эффективной популяции к переписной популяции размер был оценен как 0,6–0,7 для аутосомной и Х-хромосомной ДНК, 0,7–0,9 для митохондриальной ДНК и 0,5 для ДНК Y-хромосомы.
Эффективный размер дисперсии
Теоретические примеры
В следующих примерах одно или несколько предположений о строго идеализированной совокупности ослаблены, в то время как другие предположения сохранены. Затем рассчитывается эффективный размер дисперсии более расслабленной модели популяции относительно строгой модели.
Различия в численности населения
1 N е знак равно 1 т ∑ я знак равно 1 т 1 N я <\ displaystyle <1 \ over N_ 
Обратите внимание, что это меньше, чем среднее арифметическое размера популяции, которое в этом примере составляет 126,7. Гармоническое среднее обычно определяется наименьшим узким местом, через которое проходит популяция.
Раздельнополость
или, в более общем смысле,
где D представляет раздельнополость и может принимать значение 0 (для недельнополого) или 1 для раздельнополого.
Разница в репродуктивном успехе
Однако в естественных популяциях дисперсия часто бывает больше. Подавляющее большинство особей может не иметь потомства, а следующее поколение происходит только от небольшого числа особей, поэтому
Тогда эффективный размер популяции будет меньше и определяется по формуле:
Соотношение полов, не связанных с рыбной ловлей
Когда соотношение полов популяции варьирует от Fisherian соотношении 1: 1, эффективный размер популяции определяется по формуле:
Эффективный размер инбридинга
Для идеализированной популяции коэффициенты инбридинга подчиняются рекуррентному уравнению
Разница на поколение составляет
Эффективный размер инбридинга можно найти, решив
хотя исследователи редко используют это уравнение напрямую.
Теоретический пример: перекрывающиеся поколения и возрастные группы населения
Когда организмы живут дольше одного сезона размножения, эффективные размеры популяции должны учитывать таблицы дожития для данного вида.
Гаплоидный
Предположим гаплоидную популяцию с дискретной возрастной структурой. Примером может служить организм, способный пережить несколько отдельных сезонов размножения. Далее, определите следующие характеристики возрастной структуры:
Время генерации рассчитывается как
Тогда эффективный размер популяции инбридинга равен
Диплоид
Точно так же эффективное число инбридинга можно рассчитать для диплоидной популяции с дискретной возрастной структурой. Впервые это было дано Джонсоном, но это обозначение более похоже на Эмига и Поллака.
Предположим те же основные параметры для таблицы дожития, что и для гаплоидного случая, но с различием между мужчиной и женщиной, например N 0 ƒ и N 0 m для количества новорожденных женщин и мужчин, соответственно (обратите внимание на нижний регистр ƒ для женщин, по сравнению с заглавной буквой F для инбридинга).
Эффективное число инбридинга
Коалесцентный эффективный размер
Эффективный размер коалесцента может иметь мало отношения к количеству людей, физически присутствующих в популяции. Измеренные эффективные размеры коалесцентной популяции варьируют между генами в одной и той же популяции, будучи низкими в областях генома с низкой рекомбинацией и высокими в областях генома с высокой рекомбинацией. Время пребывания пропорционально N в нейтральной теории, но для аллелей при отборе время пребывания пропорционально log (N). Генетический автостоп может вызвать у нейтральных мутаций время пребывания, пропорциональное log (N): это может объяснить взаимосвязь между измеренным эффективным размером популяции и скоростью локальной рекомбинации.
Выбор эффективного размера
В идеализированной модели Райта-Фишера судьба аллеля, начиная с промежуточной частоты, в значительной степени определяется отбором, если коэффициент отбора s 1 / N, и в значительной степени определяется нейтральным генетическим дрейфом, если s 1 / N. В реальных популяциях значение отсечки s может зависеть от скорости локальной рекомбинации. Этот предел отбора в реальной популяции может быть зафиксирован в игрушечной симуляции Райта-Фишера посредством соответствующего выбора Ne. Прогнозируется, что в популяциях с разными эффективными размерами популяций будут развиваться совершенно разные архитектуры генома.
Эффективный размер популяции
![]()
Содержание
Обзор: Типы эффективной численности населения [ править ]
Эмпирические измерения [ править ]
В популяциях дрозофилы численностью 16 по данным переписи эффективный размер дисперсии был измерен как равный 11,5. [7] Это измерение было достигнуто путем изучения изменений частоты нейтрального аллеля от одного поколения к другому в более чем 100 реплицированных популяциях.
Для эффективной слитой численности населения, обзор публикаций по 102 в основном диким животным видов животных и растениям дало 192 Н е / N коэффициентов. В обследованных исследованиях использовалось семь различных методов оценки. Соответственно, эти отношения широко варьировались от 10 -6 для тихоокеанских устриц до 0,994 для людей, в среднем 0,34 для исследованных видов. [8] Генеалогический анализ людей-охотников-собирателей ( эскимосы) определили соотношение эффективной численности популяции к численности переписи для гаплоидного (митохондриальная ДНК, Y-хромосомная ДНК) и диплоидного (аутосомная ДНК) локусов отдельно: отношение эффективной численности популяции к переписной было оценено как 0,6–0,7 для аутосомных и Х-хромосомная ДНК, 0,7–0,9 для митохондриальной ДНК и 0,5 для Y-хромосомной ДНК. [9]
Эффективный размер дисперсии [ править ]
Теоретические примеры [ править ]
В следующих примерах одно или несколько допущений о строго идеализированной совокупности ослаблены, а другие допущения сохранены. Затем рассчитывается эффективный размер дисперсии более расслабленной модели популяции относительно строгой модели.
Различия в численности населения [ править ]
1 N е знак равно 1 т ∑ я знак равно 1 т 1 N я <\ displaystyle <1 \ over N_
Обратите внимание, что это меньше, чем среднее арифметическое размера популяции, которое в этом примере составляет 126,7. Гармоническое среднее обычно определяется наименьшим узким местом, через которое проходит популяция.
Двудомность [ править ]
или, в более общем смысле,
где D представляет раздельнополость и может принимать значение 0 (для недельнополого) или 1 для раздельнополого.
Разница в репродуктивном успехе [ править ]
Однако в естественных популяциях дисперсия часто бывает больше. Подавляющее большинство особей может не иметь потомства, а следующее поколение происходит только от небольшого числа особей, поэтому
Тогда эффективный размер популяции будет меньше и определяется по формуле:
N e ( v ) = 4 N − 2 D 2 + var ( k ) <\displaystyle N_
Соотношение полов, не связанных с рыбной ловлей [ править ]
Когда соотношение полов популяции варьирует от Fisherian соотношении 1: 1, эффективный размер популяции определяется по формуле:
N e ( v ) = N e ( F ) = 4 N m N f N m + N f <\displaystyle N_
Эффективный размер инбридинга [ править ]
Для идеализированной популяции коэффициенты инбридинга подчиняются рекуррентному уравнению
Разница на поколение составляет
Эффективный размер инбридинга можно найти, решив
N e ( F ) = 1 2 ( 1 − P t + 1 P t ) <\displaystyle N_
хотя исследователи редко используют это уравнение напрямую.
Теоретический пример: перекрывающиеся поколения и возрастные группы населения [ править ]
Когда организмы живут дольше одного сезона размножения, эффективные размеры популяции должны учитывать таблицы дожития для данного вида.
Гаплоид [ править ]
Предположим, гаплоидная популяция с дискретной возрастной структурой. Примером может служить организм, способный пережить несколько отдельных сезонов размножения. Далее, определите следующие характеристики возрастной структуры:
Время генерации рассчитывается как
Тогда эффективный размер популяции инбридинга равен [12]
Диплоид [ править ]
Точно так же эффективное число инбридинга можно рассчитать для диплоидной популяции с дискретной возрастной структурой. Впервые это было дано Джонсоном [13], но эти обозначения больше напоминают Emigh и Pollak. [14]
Предположим те же основные параметры для таблицы дожития, что и для гаплоидного случая, но с различием между мужчиной и женщиной, например, N 0 ƒ и N 0 m для количества новорожденных женщин и мужчин, соответственно (обратите внимание на нижний регистр fe для женщин, по сравнению с заглавной буквой F для инбридинга).
Эффективное число инбридинга
Коалесцентный эффективный размер [ править ]
Эффективный размер слияния может иметь мало отношения к количеству людей, физически присутствующих в популяции. [15] Измеренные эффективные размеры коалесцирующих популяций варьируются между генами в одной и той же популяции, будучи низкими в областях генома с низкой рекомбинацией и высокими в областях генома с высокой рекомбинацией. [16] [17] Время пребывания пропорционально N в нейтральной теории, но для аллелей при отборе время пребывания пропорционально log (N). Генетический автостоп может привести к тому, что нейтральные мутации имеют время пребывания, пропорциональное log (N): это может объяснить взаимосвязь между измеренным эффективным размером популяции и скоростью локальной рекомбинации.
Эффективный размер выделения [ править ]
В идеализированной модели Райта-Фишера судьба аллеля, начиная с промежуточной частоты, в значительной степени определяется отбором, если коэффициент отбора s 1 / N, и в значительной степени определяется нейтральным генетическим дрейфом, если s 1 / N. В реальных популяциях значение отсечки s может зависеть от скорости локальной рекомбинации. [6] [18] Этот предел отбора в реальной популяции может быть зафиксирован в игрушечном моделировании Райта-Фишера посредством соответствующего выбора Ne. Прогнозируется, что в популяциях с разными эффективными размерами популяций будут развиваться совершенно разные архитектуры генома. [19] [20]
6. Популяция и ее структура
М.Э. Гусельников, В.Н. Стройнова
Биоэкология
Учебное пособие. – Томск: Изд. ТПУ, 2002. – 104 с.
6. Популяция и ее структура
Популяция и ее законы
Что такое популяция и чем она отличается от биологического вида?
Вид — это группа особей, обладающих общими признаками. Эти признаки следующие:
1. Строение тела, физиология и способы взаимоотношений со средой.
2. Способность к скрещиванию и образованию плодовитого потомства.
3. Особи одного вида населяют определенную территорию – АРЕАЛ.
4. Особи одного вида отличаются от других групп особей практически полным отсутствием гибридных форм. То есть один вид не скрещивается с другим в пределах одного отряда.
Однако особи одного вида распределены в пределах ареала неравномерно. Существуют участки с высокой численностью особей вида, средней численностью в соседней местности, с низкой численностью. Эти центры плотности населения вида являются популяциями. Главная функция популяции в том, что только популяция может воспроизводить новые поколения вида в условиях конкретной экосистемы. В то же время особи близких видов не могут скрещиваться между собой. Популяция, являясь частью сообщества, функционирует как его часть. Экологические взаимоотношения в сообществе и экосистеме определяются не видом, а популяцией. Таким образом, популяция представляет собой открытую систему, осуществляющую обмен энергией, веществом и информацией с окружающей средой (экосистемой). Внутри популяции также существуют потоки энергии, информации и вещества. Основными количественными характеристиками популяций являются численность и плотность. Численность — это число отдельных особей в популяции, плотность — число особей на единицу занимаемой ими площади или объема.
Особи одной популяции отличаются от особей близкой популяции, например, внешними признаками, в соответствии с биотопом, в котором обитают. Например, белки штата Вашингтон серые круглый год, а наши сибирские белки рыжевато-бурые летом, серовато-бурые зимой.
Таким образом, популяция представляет собой незамкнутую систему, взаимодействующую с внешней средой. Вспомните законы термодинамики и энтропии, которые мы рассматривали при изучении экосистем. Системные законы справедливы и для популяции. Рассмотрим также некоторые экологические законы, действующие внутри популяции и определяющие ее динамику (развитие).
1. ПРИНЦИП МИНИМАЛЬНОГО И МАКСИМАЛЬНОГО РАЗМЕРА ПОПУЛЯЦИИ: размер популяции не может быть меньше минимума, который необходим для ее стабильности во внешних условиях и воспроизводства. Размер популяции не может быть больше максимума, что определяется пищевыми ресурсами и их доступностью, а также условиями размножения. Числа минимума и максимума различны для разных популяций. Внутри популяции колебания численности наблюдаются между этими двумя числами.
2. ПРАВИЛО КОЛЕБАНИЙ ЧИСЛЕННОСТИ: никакая популяция не может находиться в состоянии абсолютной уравновешенности числа особей. Обязательно возникают периодические осцилляции, обусловленные внешними (приток энергии, пищевые ресурсы) или внутренними факторами.
3. ПРИНЦИП НИКОЛСОНА: популяции есть стабильные системы, способные противостоять факторам внешней среды и контролировать их изменением плотности популяции.
4. ПРИНЦИП ВОЗДЕЙСТВИЯ ФАКТОРОВ: границы, размер и характер места обитания популяции обусловлены их биологическими особенностями. Наоборот, биологические особенности популяции являются индикатором места обитания.
Понятие популяции применимо как к животным, так и к растениям. Любые популяции имеют определенную пространственную, возрастную, половую и генетическую структуры.
1. Пространственная структура популяции определяет эффективное использование ресурсов среды (пища) и снижает конкуренцию внутри популяции. Тип использования территории может быть двояким и определяется образом жизни. При оседлом образе жизни используется одна и та же территория в течение жизни нескольких поколений, при кочевом образе жизни используются различные территории внутри ареала вида. Пространственная структура популяции поддерживается агрессивными методами поведения, чужаки изгоняются с территории данной популяции.
Интересна пространственная структура популяций растений (рис. 6.1), она имеет вертикальную (ярусность леса) и горизонтальную составляющие.
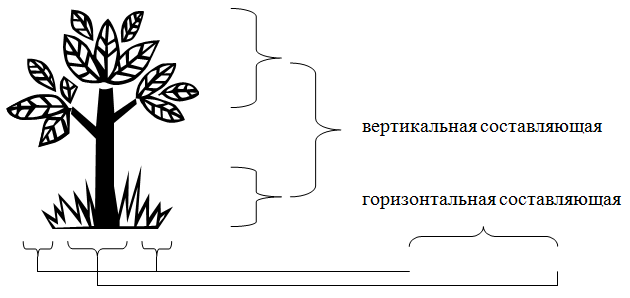
Рис. 6.1. Пространственная структура популяций
2. В популяциях животных или растений имеются различные возрастные группы особей. В популяциях высших животных, например человека, возраст молодости, зрелости, старости составляет примерно 1/3 от продолжительности жизни. Такое соотношение между возрастными группами обеспечивает устойчивое воспроизводство популяции. Доминирование возрастной группы определяет возраст популяции. Возраст популяции зависит от рождаемости, смертности и выживаемости. Рождаемость К — это число рождений в единицу времени, смертность Р — число смертей в единицу времени. Выживаемость Р / К — средняя для популяции вероятность сохранения особей каждого поколения за определенный промежуток времени. В молодых популяциях рождаемость превышает смертность, в старых — наоборот. Кривая выживания характеризует популяцию, она строго индивидуальна. На рис. 6.2 представлена кривая выживания для различных организмов.
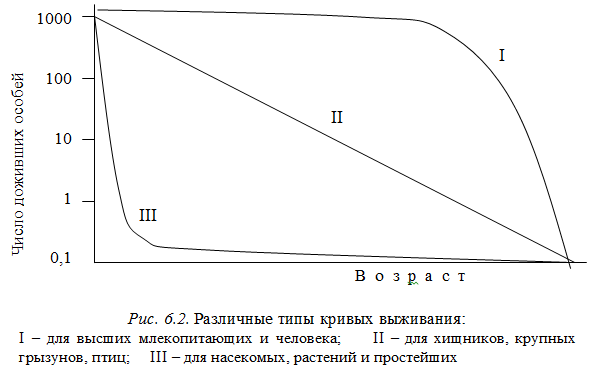
Кривая типа I наблюдается у высших животных и человека, K > P. В этом случае особи живут долго и умирают от старости. Кривая типа II характерна для хищников, крупных грызунов, птиц, когда К=Р. При этом наблюдается равновесие между рождаемостью и смертностью, до старости доживают лишь отдельные особи. Кривая типа III наблюдается у насекомых, рыб, простейших. К



