Конъюгация хромосом
Мейоз (или редукционное деление клетки) — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток, или гамет, из недифференцированных стволовых.
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса.
В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма.
Этот же механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет. Определенные ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации).
Фазы мейоза
Мейоз состоит из двух последовательных делений с короткой интерфазой между ними.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и два так называемых редукционных тельца (абортивные дериваты первого и второго делений).
КОНЪЮГАЦИЯ ХРОМОСОМ
КОНЪЮГАЦИЯ ХРОМОСОМ (лат. conjugatio соединение; хромосомы; син.: синапсис хромосом, спаривание хромосом) — тесное соединение хромосом друг с другом у всех организмов, включая человека, обладающих сформированным клеточным ядром.

Различают конъюгацию гомологичных и негомологичных хромосом. Конъюгация гомологичных хромосом является обязательным этапом мейоза (см.), а также происходит в некоторых соматических клетках, напр, при формировании политенных (гигантских) хромосом (см.) в клетках слюнных желез личинок мух, комаров и других двукрылых насекомых. Этот тип К. х. отличается тем, что гомологичные хромосомы за счет специфического взаимного «узнавания» и притяжения гомологичных генов плотно прилегают одна к другой по всей длине так, что хромомеры, содержащие гомологичные (аллельные) гены, находятся точно друг против друга. Если в одной из конъюгирующих хромосом произошла транслокация (см.) или инверсия (см.), то участок хромосомы с измененной последовательностью расположения генов не способен конъюгировать с противолежащим участком второй, гомологичной хромосомы (рис. 1). Однако если перестройка произошла на достаточно длинном отрезке, то хромосомы-партнеры, образуя петлю или крест, способны обеспечить сопоставление гомологичных локусов и тем самым осуществить конъюгацию.
В политенных хромосомах конъюгация гомологов носит необратимый характер, гомологичные хромосомы остаются соединенными до конца существования слюнных желез. Биол. значение такой К. х. остается неясным. Конъюгация гомологичных хромосом в мейозе носит обратимый характер и лежит в основе точного разделения диплоидного набора хромосом на два гаплоидных набора, которые расходятся в разные клетки (редукция числа хромосом). Это явление создает условия для полового процесса и генетической рекомбинации у диплоидных организмов. Процесс конъюгации гомологичных хромосом происходит В профазе I мейотического деления и начинается на стадии зиготены. При этом сначала происходит сближение гомологичных хромосом с расстояния нескольких микрометров до расстояния примерно 0,2 мкм, обеспечивающего контакт хромосом. Затем происходит собственно «узнавание» и специфическое притяжение гомологичных хромомеров (локусов генов). В результате две конъюгирующие гомологичные хромосомы ложатся параллельно одна другой так, что гомологичные хромомеры образуют пары. Две соединенные гомологичные хромосомы носят название бивалента, а стадия мейоза, на к-рой соединение (конъюгация) завершилось по всей длине хромосом, называется пахитеной. Во время пахитены происходит кроссинговер (перекрест) — обмен участками гомологичных хромосом (см. Рекомбинация). На стадии пахитены каждая хромосома состоит из двух продольных половин — хроматид (4 хроматиды в биваленте). В данном месте бивалента кроссинговер происходит только между двумя несестринскими хроматидами из четырех. На следующей стадии — в диплотене — гомологичные хромосомы отталкиваются одна от другой во всех точках бивалента, кроме тех, где произошли перекресты. В результате места перекрестов (хиазмы) становятся видимыми в микроскоп. На следующих стадиях мейоза — в диплотене, диакинезе и метафазе I деления под влиянием конденсации и укорочения хромосом хиазмы перемещаются к концам бивалентов. Происходит так наз. терминализация хиазм. При этом хиазмы продолжают удерживать пары бывших партнеров по конъюгации. В метафазе I каждая хромосома в биваленте соединяется нитью веретена только с одним полюсом клеточного деления. Благодаря этому в анафазе I мейотического деления гомологичные хромосомы расходятся к противоположным полюсам и к каждому полюсу попадает по одной хромосоме из каждого бивалента. Таким образом, К. х. и хиазмы обеспечивают правильную редукцию числа хромосом. Если в кариотипе (см.) есть непарные хромосомы, напр, единичная половая хромосома в норме у самцов некоторых видов насекомых или одиночная половая хромосома у человека при синдроме Тернера, то такие хромосомы не вступают в конъюгацию из-за отсутствия партнера, остаются унивалентными в профазе I и случайным образом направляются к тому или иному полюсу в анафазе I. У противоположного полюса оказывается набор, лишенный одной хромосомы. У межвидовых гибридов, напр, у мула, нет ни одной парной хромосомы, т. к. половина хромосом получена от хромосомного набора лошади, а другая — осла. В результате в профазе I К. х. вообще отсутствует, и все хромосомы остаются унивалентными. В анафазе I деления они хаотически расходятся к полюсам, и дочерние клетки, а также формирующиеся из них гаметы получают несбалансированные как по видовому составу, так и по числу наборы хромосом. Это приводит к нежизнеспособности гамет или зигот. Бесплодие гибридов типа мула обусловлено отсутствием у них К. х. в мейозе.
Появление в диплоидном хромосомном наборе (см.) лишней хромосомы (трисомия по данной хромосоме) приводит к нарушению К. х. из-за конкуренции между тремя партнерами по конъюгации. Исследование таких случаев привело к открытию правила, согласно к-рому в каждой точке (локусе) хромосомы возможна конъюгация только двух партнеров. Однако в другом локусе может произойти смена партнера, и в результате появляются триваленты. Это также нарушает расхождение хромосом в анафазе I. Различные случаи нарушения К. х. приводят к появлению гипогаплоидных или гипергаплоидных гамет (недостаток хромосом или их избыток). Если такие гаметы выживают и формируют зиготу, то возникают анэуплоидные зародыши (нарушение строгой диплоидности), при этом возможны случаи моносомии (отсутствие одной из хромосом), трисомии (появление лишней хромосомы) и т. п. У человека моно- и трисомия по половым хромосомам и нек-рым аутосомам приводит к хромосомным болезням (см.), а анэуплоидия по крупным аутосомам — к гибели плода во внутриутробном периоде.
Иногда у гибридов близких видов, в частности у растительных гибридов, или у организмов, несущих крупные перестройки хромосом, наблюдается частичная гомология хромосом. Тогда о способности хромосом к конъюгации судят по числу бивалентов в клетке на стадиях диплотены — метафазы I и о «силе» этой конъюгации — по числу хиазм на бивалент. Однако известны случаи ахиазматии (полного отсутствия хиазм) при нормально протекающей конъюгации хромосом в зиготене и пахитене. Преждевременное разъединение гомологов (распад бивалентов) из-за отсутствия хиазм называют десинапсисом. Десинапсис приводит к такому же нарушению расхождения хромосом в анафазе I, какое наблюдается у мула.

Установлено, что процесс конъюгации гомологичных хромосом в мейозе находится под контролем многих генов, действующих только во время мейоза. Такие гены обнаружены у дрозофилы, грибов и ряда высших растений, но их действие оказывается одинаковым у самых разных организмов. Генетический контроль К. х. свидетельствует о том, что К. х. обеспечивается синтезируемыми во время мейоза специфическими белками. Сближение хромосом с дальних расстояний, вероятно, осуществляется за счет каких-то факторов в ядерной мембране: концы гомологичных хромосом, прикрепленные к ядерной мембране, «скользят» по ней навстречу друг другу, обеспечивая тем самым сближение хромосом. Возможно, что сближение хромосом с дальних расстояний происходит за счет неспецифического взаимодействия ДНК, локализованной в прицентромерных участках хромосом, за счет интеркалярного гетерохроматина (т. е. гетерохроматина, находящегося между двумя дисками эухроматина), способного к так наз. эктопическому спариванию — временной конъюгации негомологичных участков хромосом. Более известны молекулярные механизмы взаимодействия хромосом на близких расстояниях. Доказано участие в этом процессе уникальных последовательностей нуклеотидов ДНК, расположенных по ли локально по длине всех хромосом и реплицирующихся накануне или во время К. х. на стадии зиготены (зетДНК). Установлено, что К. х. на стадии зиготены сопровождается формированием так наз. синаптонемального комплекса (СК). Он формируется в процессе К. х. в мейозе у всех эукариотических организмов (от инфузорий и дрожжей до человека) и представляет собой субмикроскопическую структуру, к-рая состоит из трех рибонуклеопротеидных тяжей, идущих вдоль каждой пары гомологичных хромосом внутри бивалента (рис. 2). Два наружных тяжа называются боковыми элементами СК, внутренний — центральным. Боковые элементы возникают в каждой хромосоме до их конъюгации и сближаются в момент конъюгации до расстояния 150—200 нм. В это время между ними формируется центральный элемент. Предполагается, что центральный элемент служит местом формирования гетеро-дуплексов зетДНК (гибридных молекул ДНК), в к-рой полинуклеотидные нити двойной спирали принадлежат разным хромосомам — партнерам в биваленте. Предполагается, что СК препятствует необратимому соединению гомологичных хромосом, удерживая их на строго определенном расстоянии, ибо он отторгается от хромосом после прекращения их конъюгации на стадии диплотены, разрушается и выводится из ядра нередко в виде поликомплексов, упакованных в трехмерный штабель отрезков боковых и центральных элементов. СК отсутствует при необратимой К. х. в политенных хромосомах. Доказано, что формирование СК является результатом активации в мейозе специфических генов, в частности нормального аллеля гена с (3) G у дрозофилы. Формирование СК обеспечивает высокую частоту кроссинговера, но не является непременным условием для его осуществления; в отсутствие СК кроссинговер может идти, но с пониженной частотой.
Конъюгация негомологичных хромосом (конъюгация гомологичных участков в негомологичных хромосомах) наблюдается в мейозе у гаплоидных растений, во время митоза (см.) в соматических клетках многих растений и животных (эктопическое спаривание). На основе неслучайного расхождения негомологичных хромосом в мейозе у дрозофил с перестроенным хромосомным набором можно сделать вывод, что в профазе I мейоза происходила конъюгация негомологичных хромосом. Экспериментально подтверждается гипотеза о полилокальном расположении в хромосомах дрозофилы определенной фракции ДНК, состоящей из так наз. умеренных повторов нуклеотидов. Они способны обеспечить взаимное «узнавание» идентичных участков в той же самой и других хромосомах клетки и обусловить, т. о., конъюгацию негомологичных хромосом.
Митоз и мейоз
Жизненный цикл клетки (клеточный цикл)
С момента появления клетки и до ее смерти в результате апоптоза (программируемой клеточной гибели) непрерывно продолжается жизненный цикл клетки.
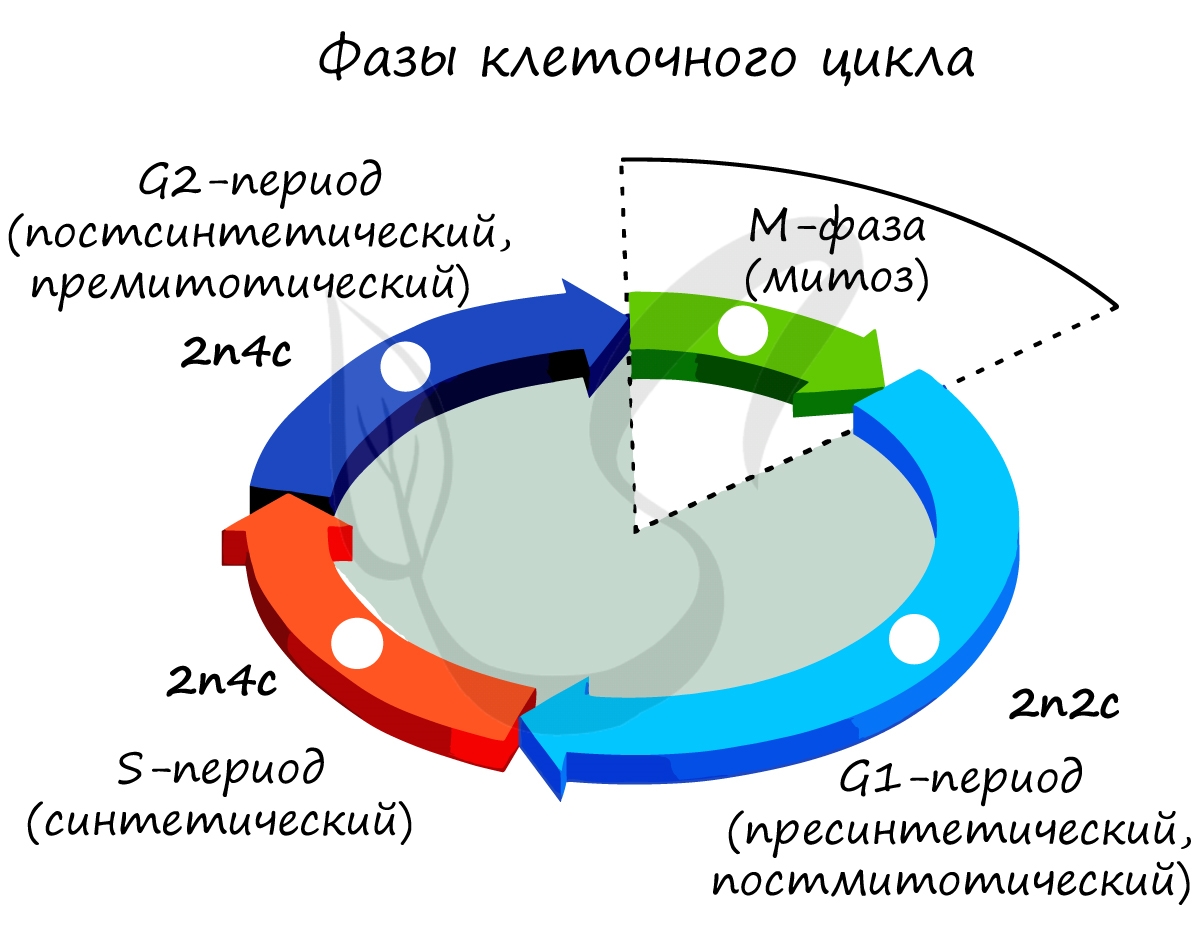
Интенсивно образуются рибосомы, синтезируется АТФ и все виды РНК, ферменты, клетка растет.
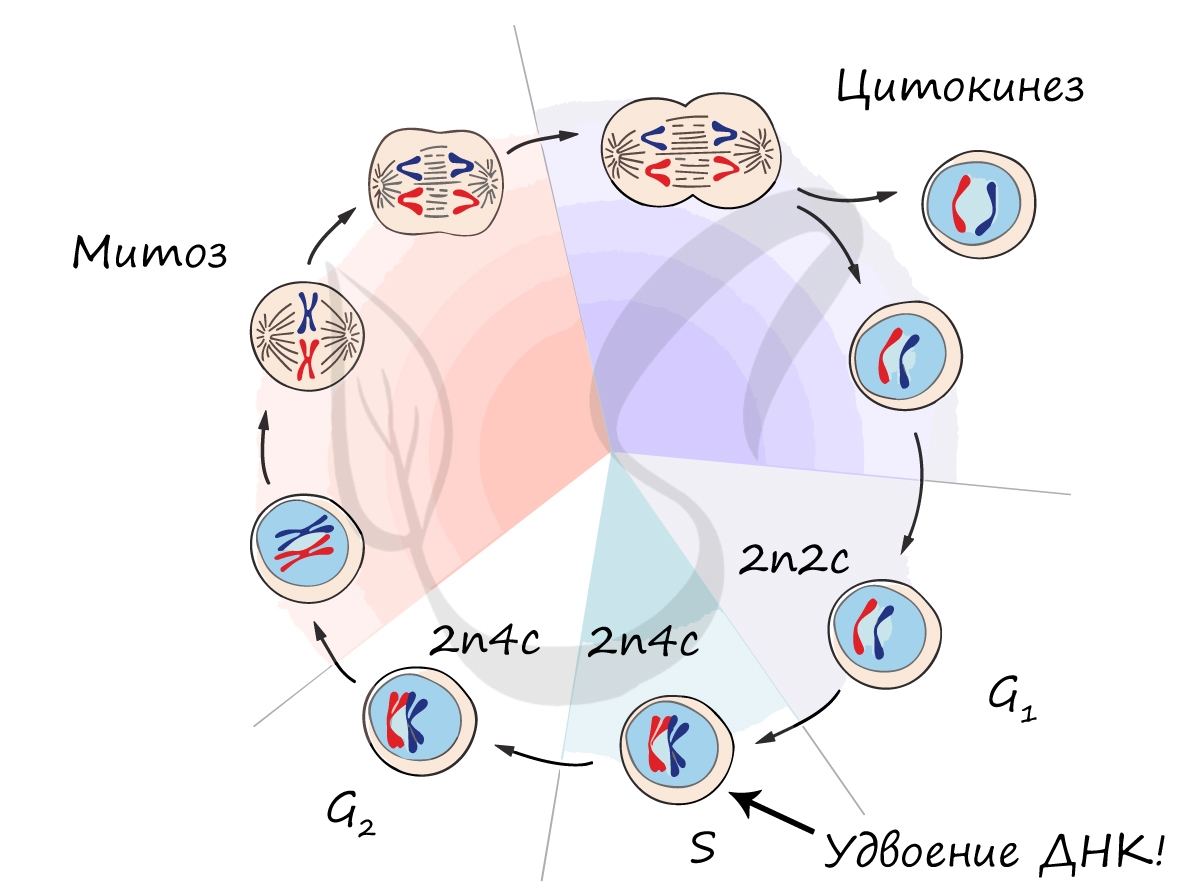
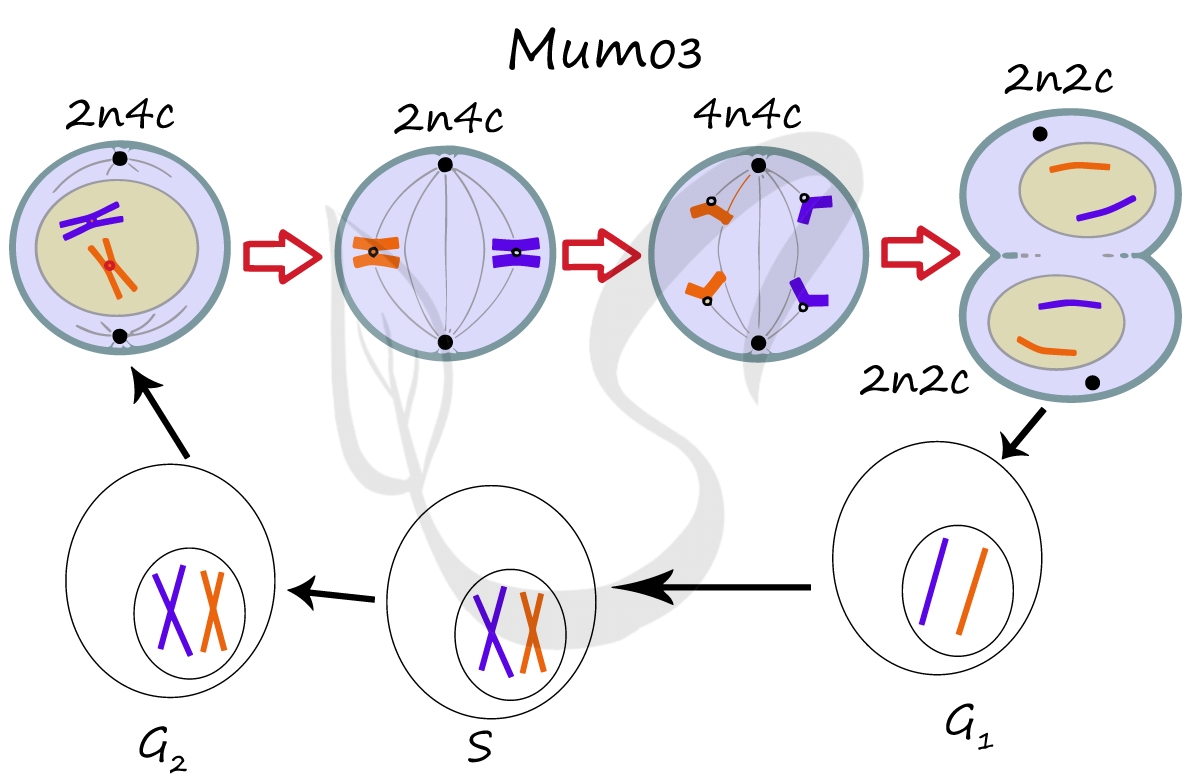
Митоз является непрямым способом деления клетки, наиболее распространенным среди эукариотических организмов. По продолжительности занимает около 1 часа. К митозу клетка готовится в период интерфазы путем синтеза белков, АТФ и удвоения молекулы ДНК в синтетическом периоде.
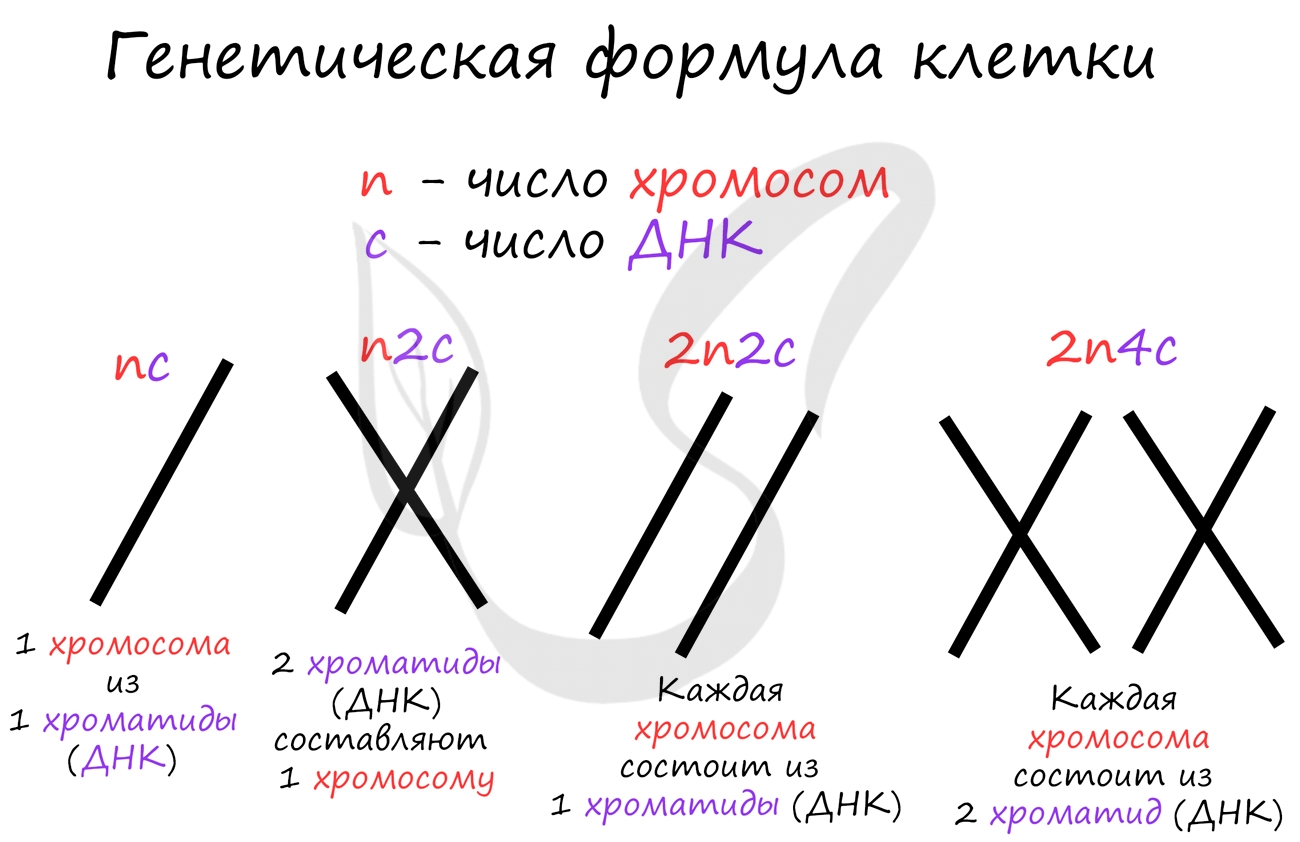
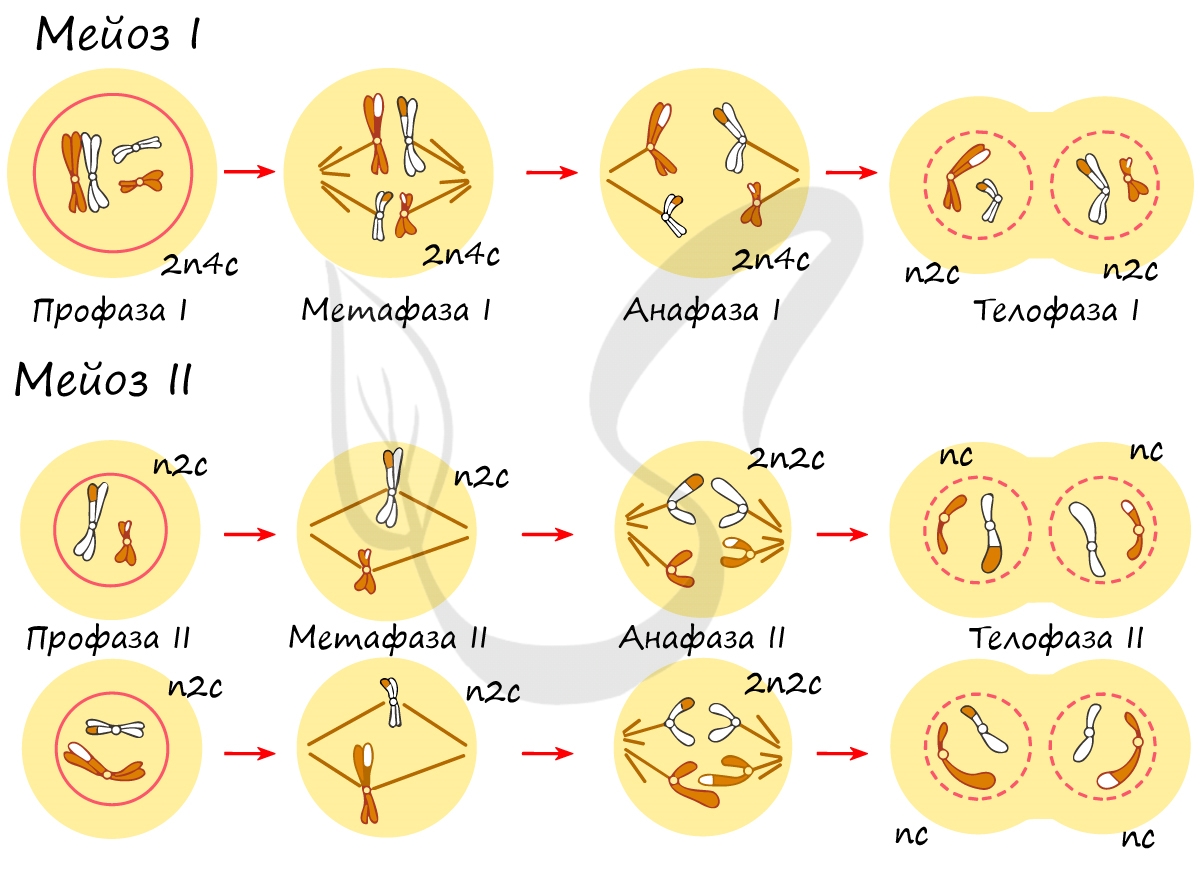
Митоз состоит из 4 фаз, которые мы далее детально рассмотрим: профаза, метафаза, анафаза, телофаза. Напомню, что клетка вступает в митоз с уже удвоенным (в синтетическом периоде) количеством ДНК. Мы рассмотрим митоз на примере клетки с набором хромосом и ДНК 2n4c.
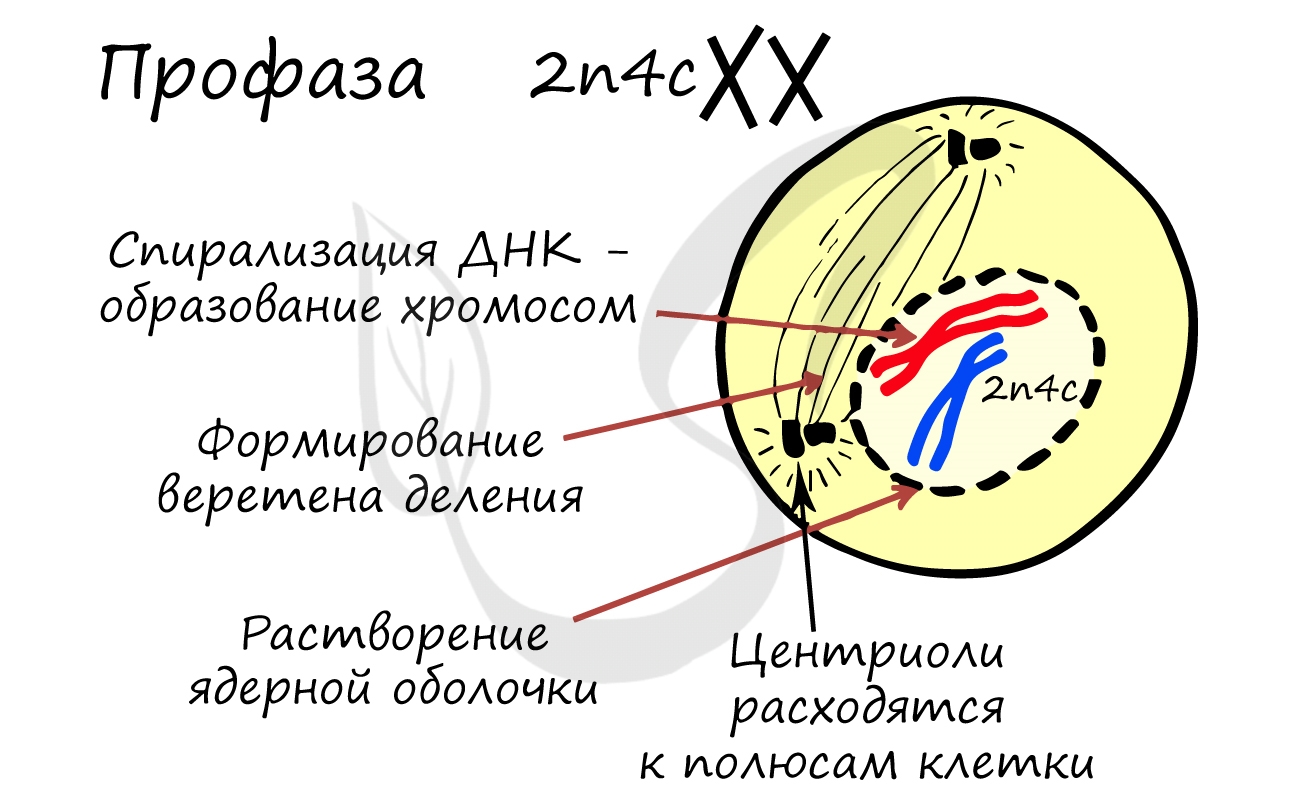
ДНК максимально спирализована в хромосомы, которые располагаются на экваторе клетки. Каждая хромосома состоит из двух хроматид, соединенных центромерой (кинетохором). Нити веретена деления прикрепляются к центромерам хромосом (если точнее, прикрепляются к кинетохору центромеры).
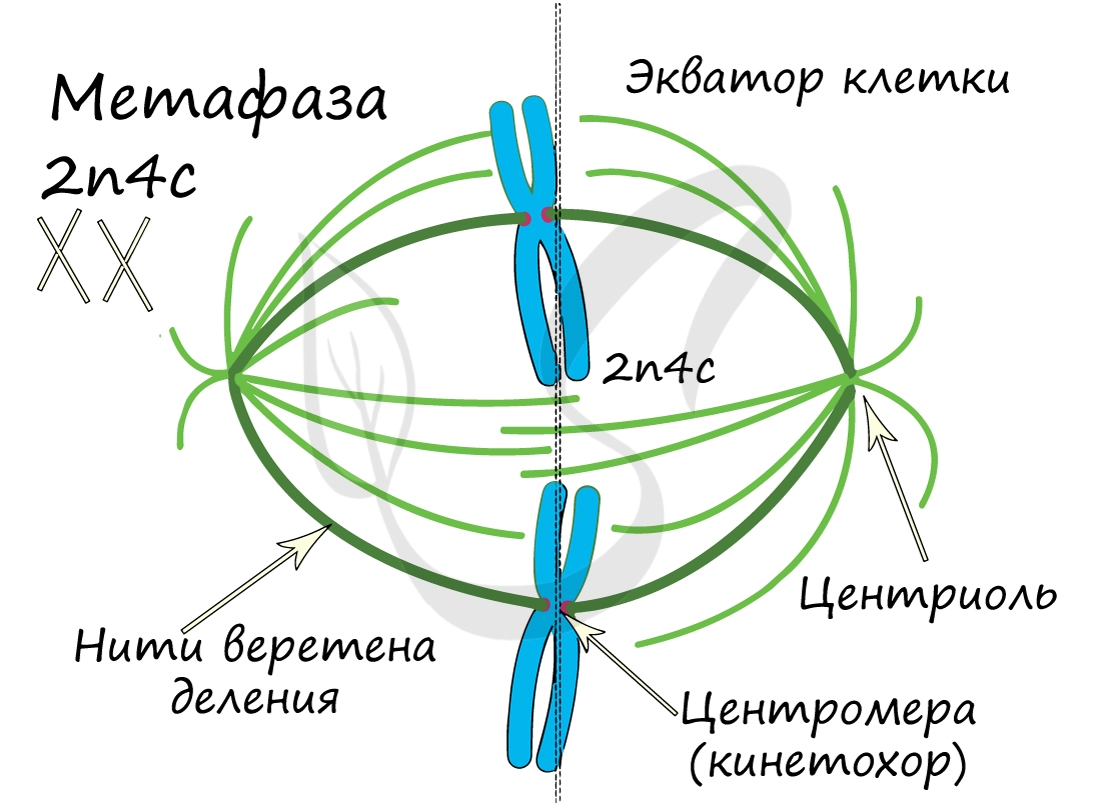
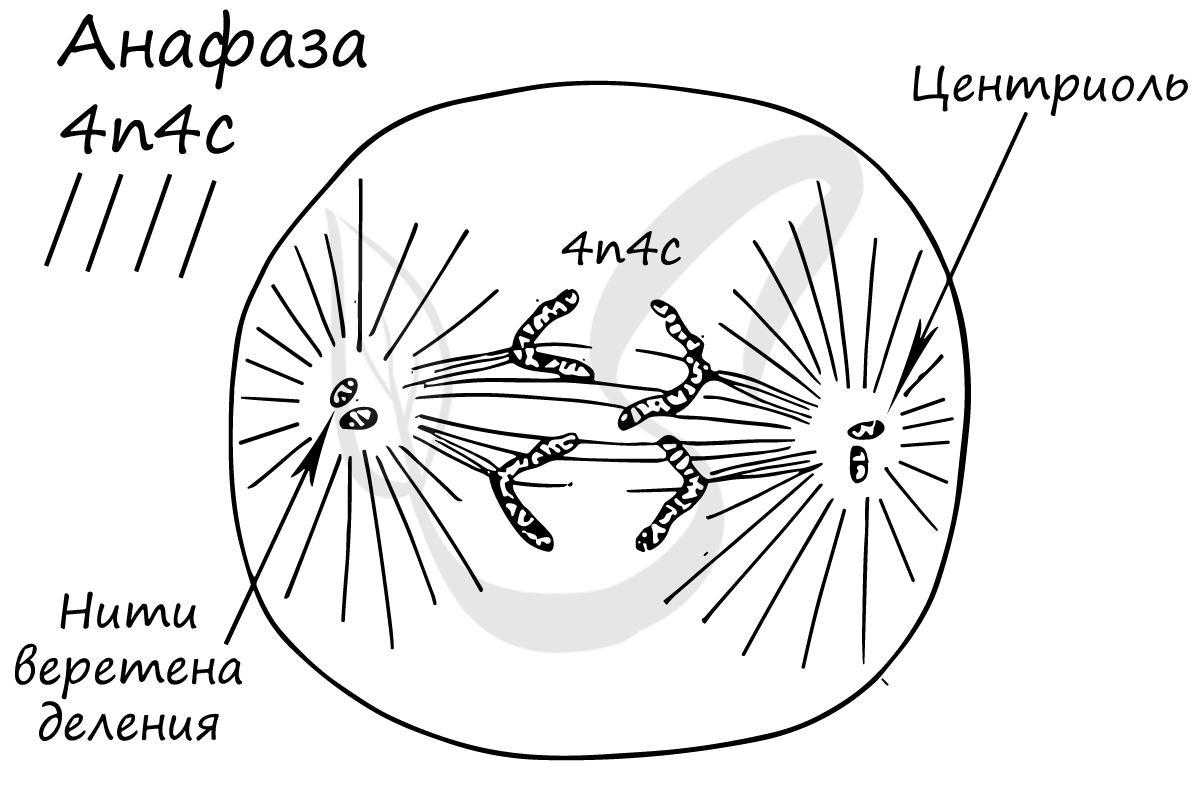
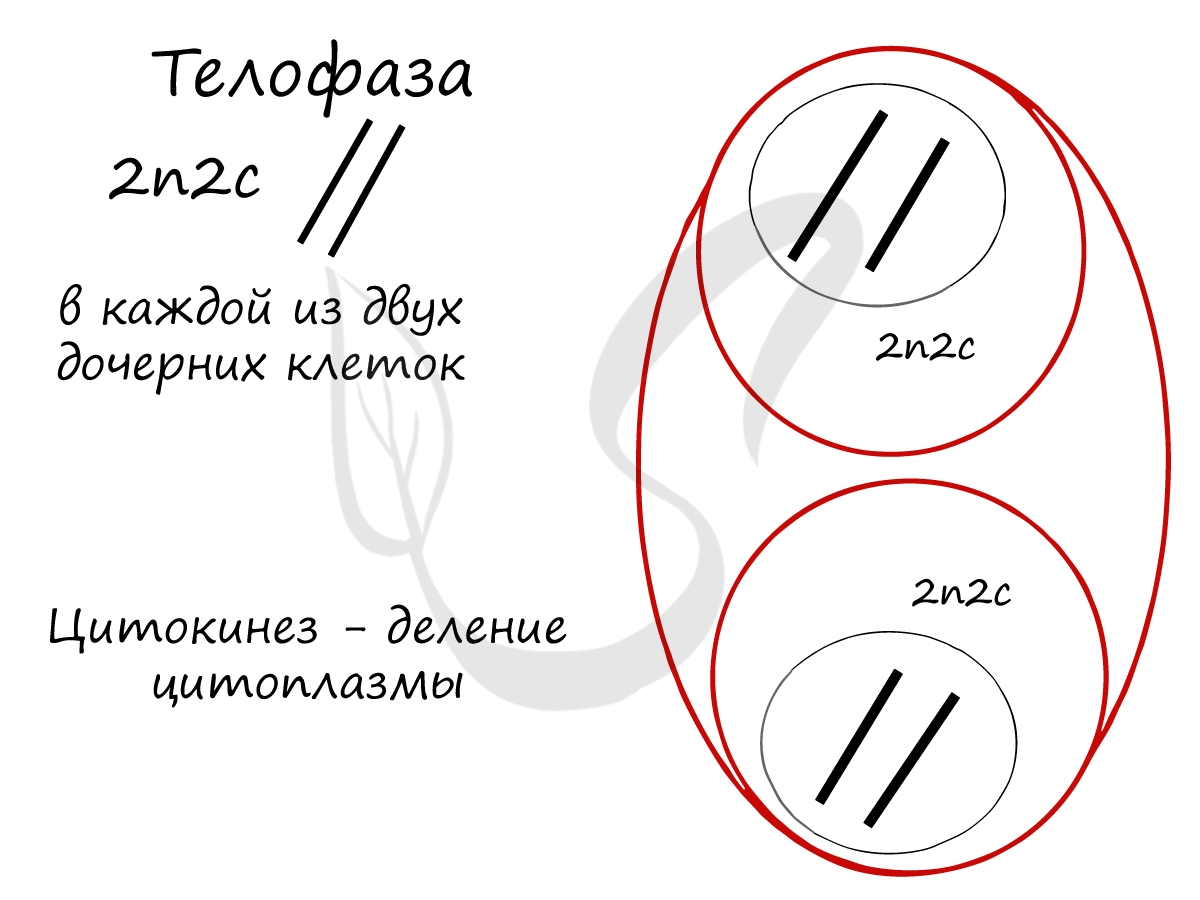
Попробуйте самостоятельно вспомнить фазы митоза и описать события, которые в них происходят. Особенное внимание уделите состоянию хромосом, подчеркните сколько в них содержится молекул ДНК (хроматид).
Мейоз
В результате мейоза из диплоидных клеток (2n) получаются гаплоидные (n). Мейоз состоит из двух последовательных делений, между которыми практически отсутствует пауза. Удвоение ДНК перед мейозом происходит в синтетическом периоде интерфазы (как и при митозе).
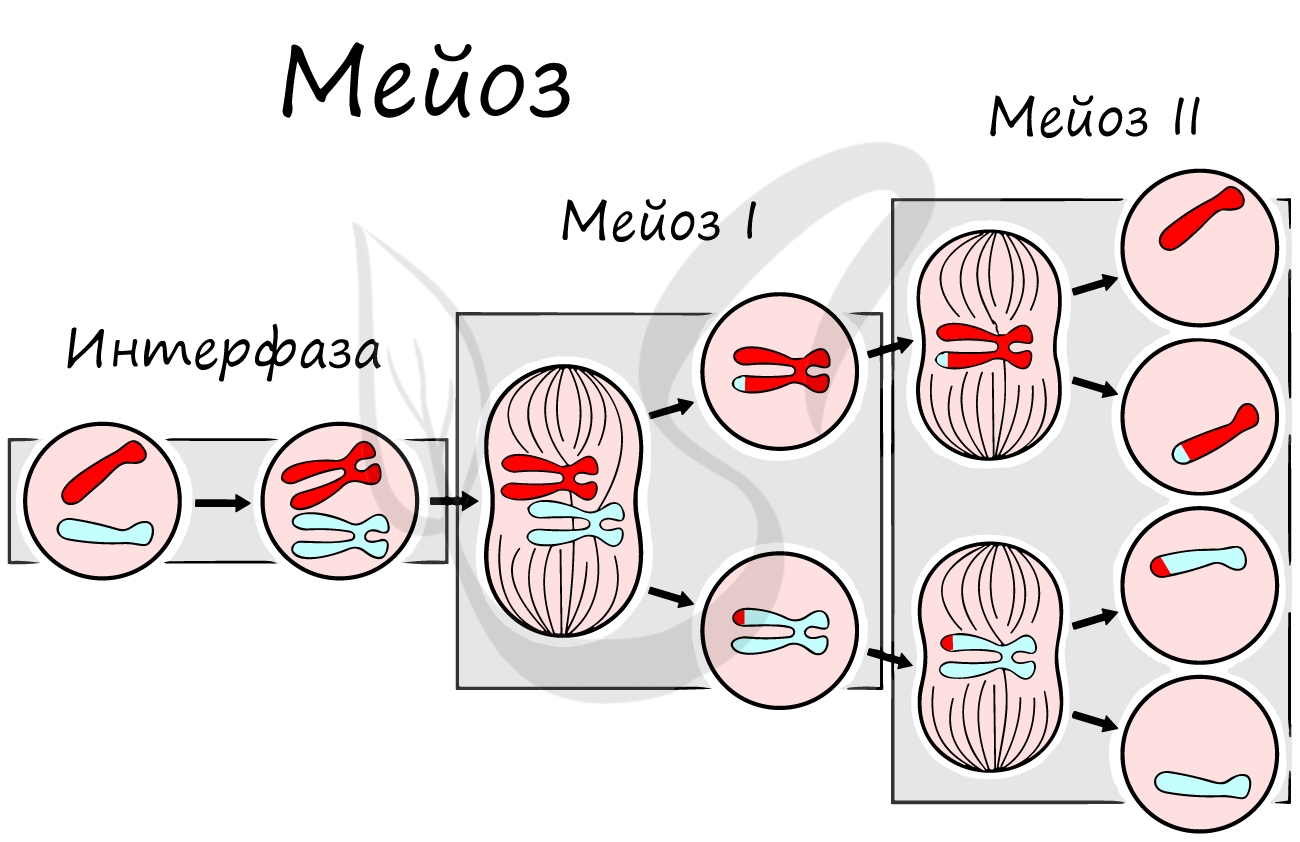
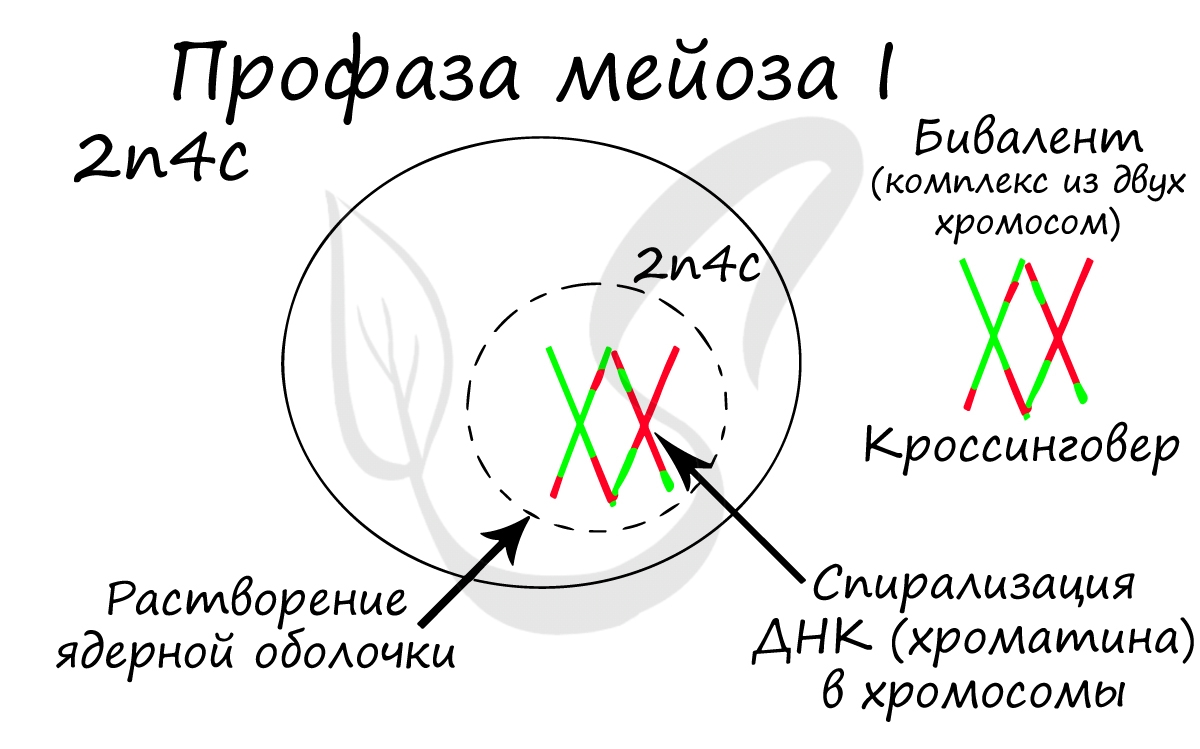
Помимо типичных для профазы процессов (спирализация ДНК в хромосомы, разрушение ядерной оболочки, движение центриолей к полюсам клетки) в профазе мейоза I происходят два важнейших процесса: конъюгация и кроссинговер.
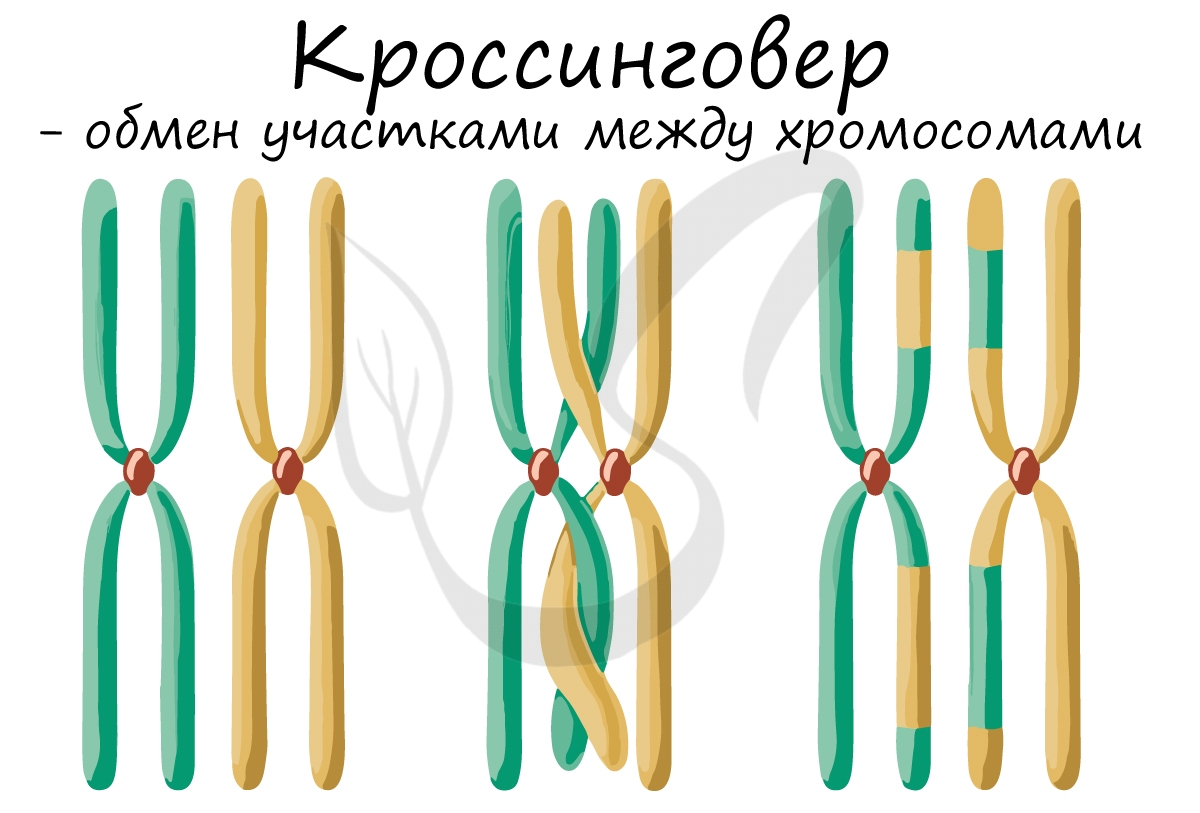
Кроссинговер является важнейшим процессом, в ходе которого возникают рекомбинации генов, что создает уникальный материал для эволюции, последующего естественного отбора. Кроссинговер приводит к генетическому разнообразию потомства.
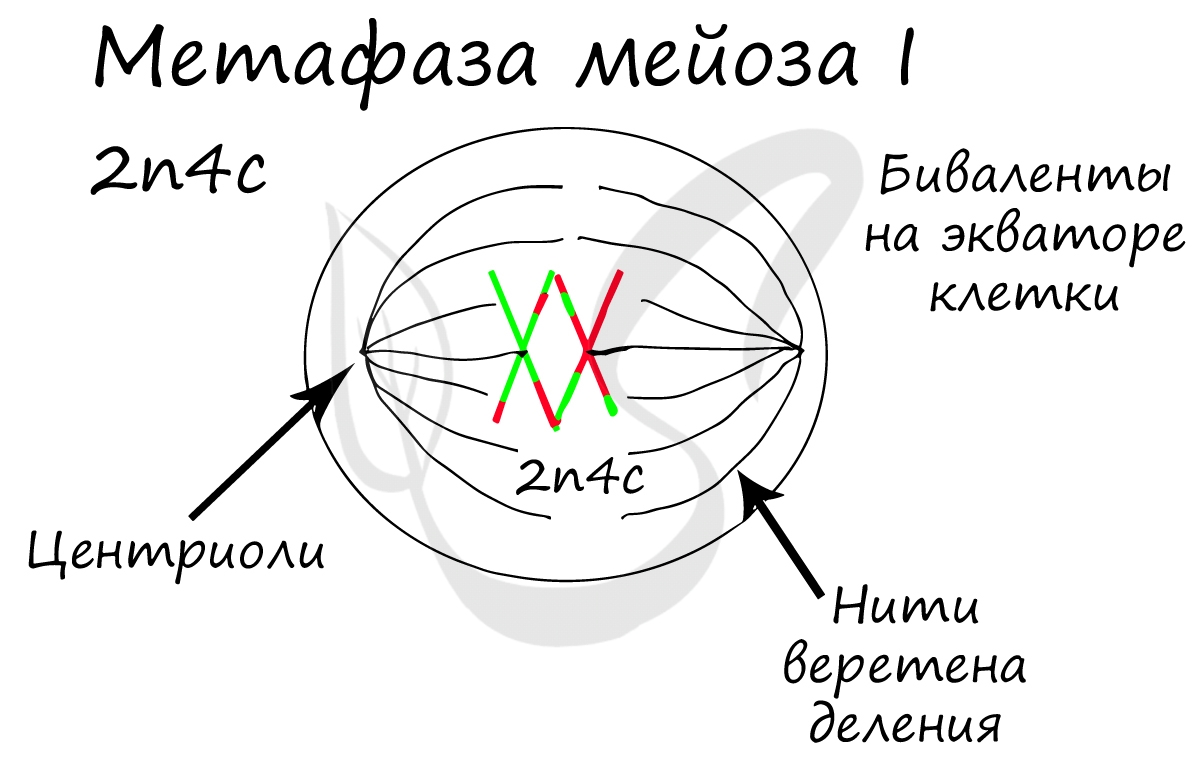
Биваленты (комплексы из двух хромосом) выстраиваются по экватору клетки. Формируется веретено деления, нити которого крепятся к центромере (кинетохору) каждой хромосомы, составляющей бивалент.

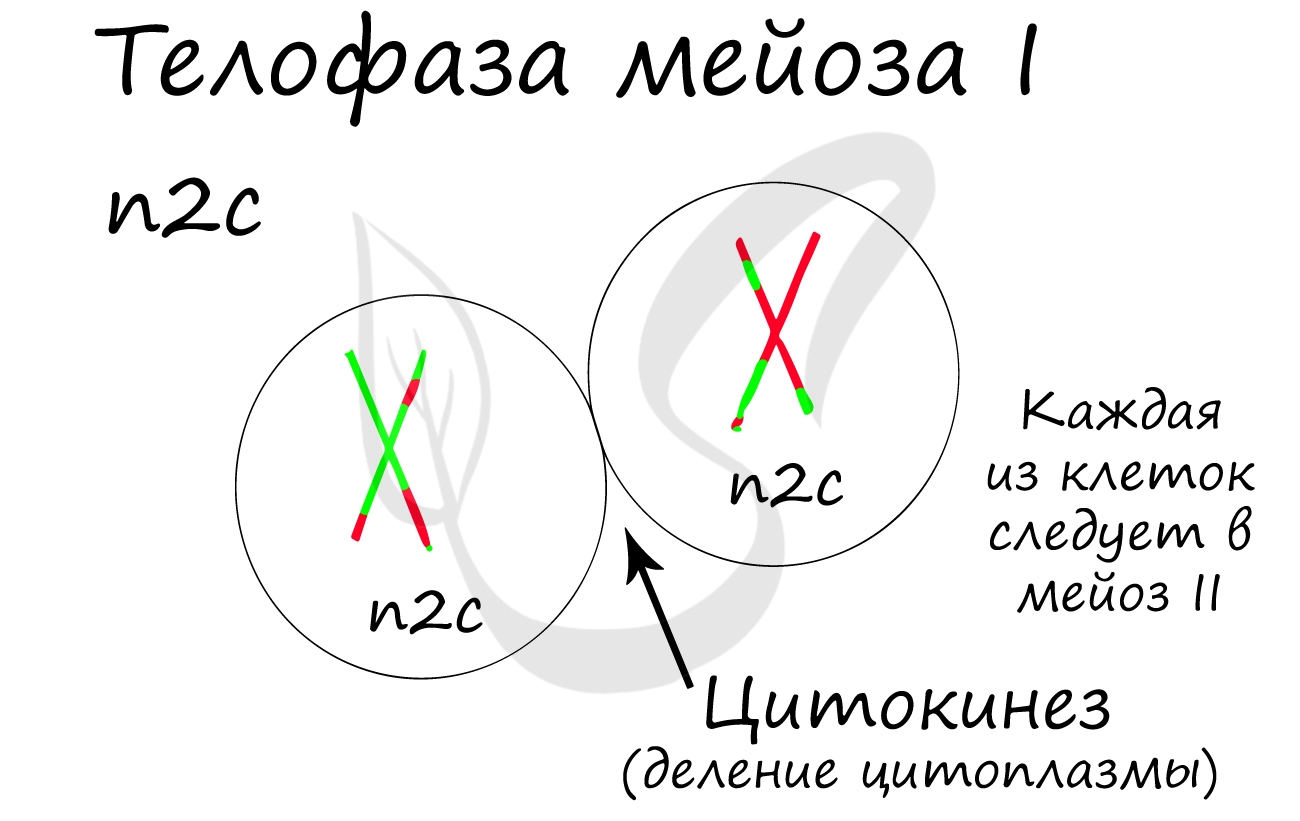
Мейоз II весьма напоминает митоз по всем фазам, поэтому если вы что-то подзабыли: поищите в теме про митоз. Главное отличие мейоза II от мейоза I в том, что в анафазе мейоза II к полюсам клетки расходятся не хромосомы, а хроматиды (дочерние хромосомы).
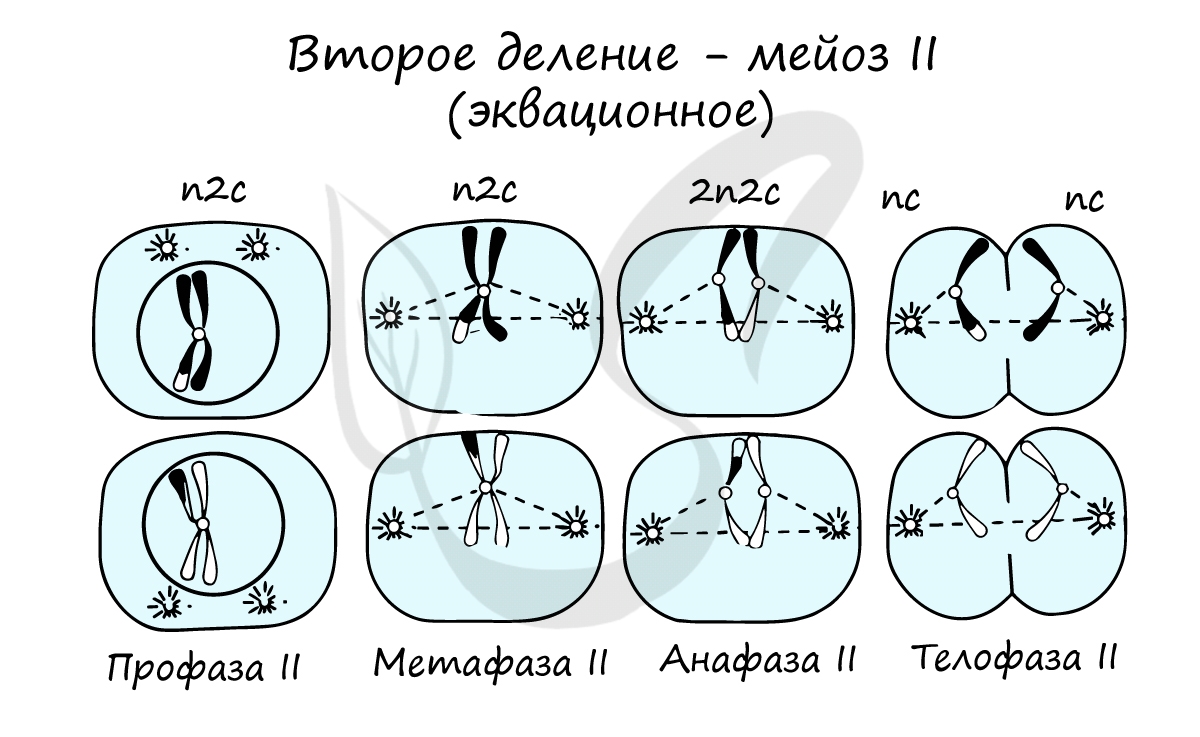
Сейчас мы возьмем клетку, в которой 4 хромосомы. Попытайтесь самостоятельно описать фазы и этапы, через которые она пройдет в ходе мейоза. Проговорите и осмыслите набор хромосом в каждой фазе.
Бинарное деление надвое
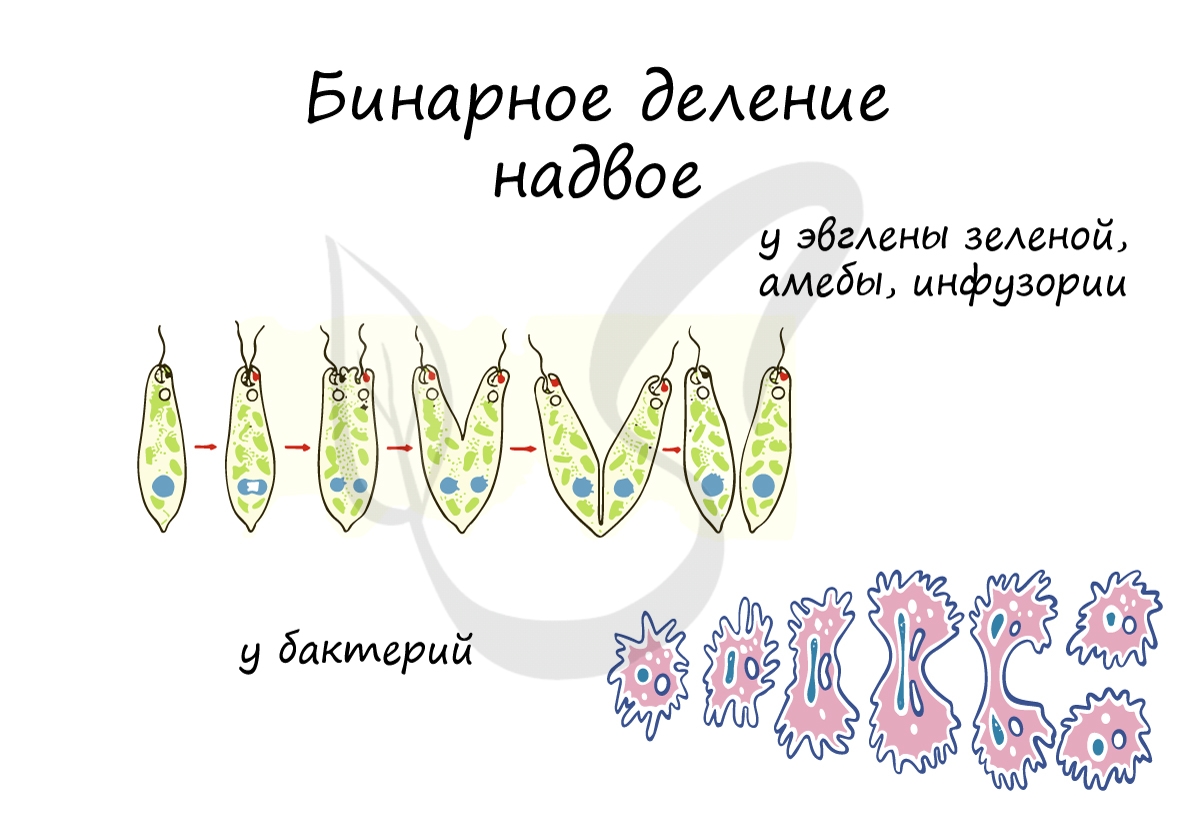
При благоприятных условиях бактерии делятся каждые 20 минут. В случае, если условия не столь благоприятны, то больше времени уходит на рост и развитие, накопление питательных веществ. Интервалы между делениями становятся длиннее.
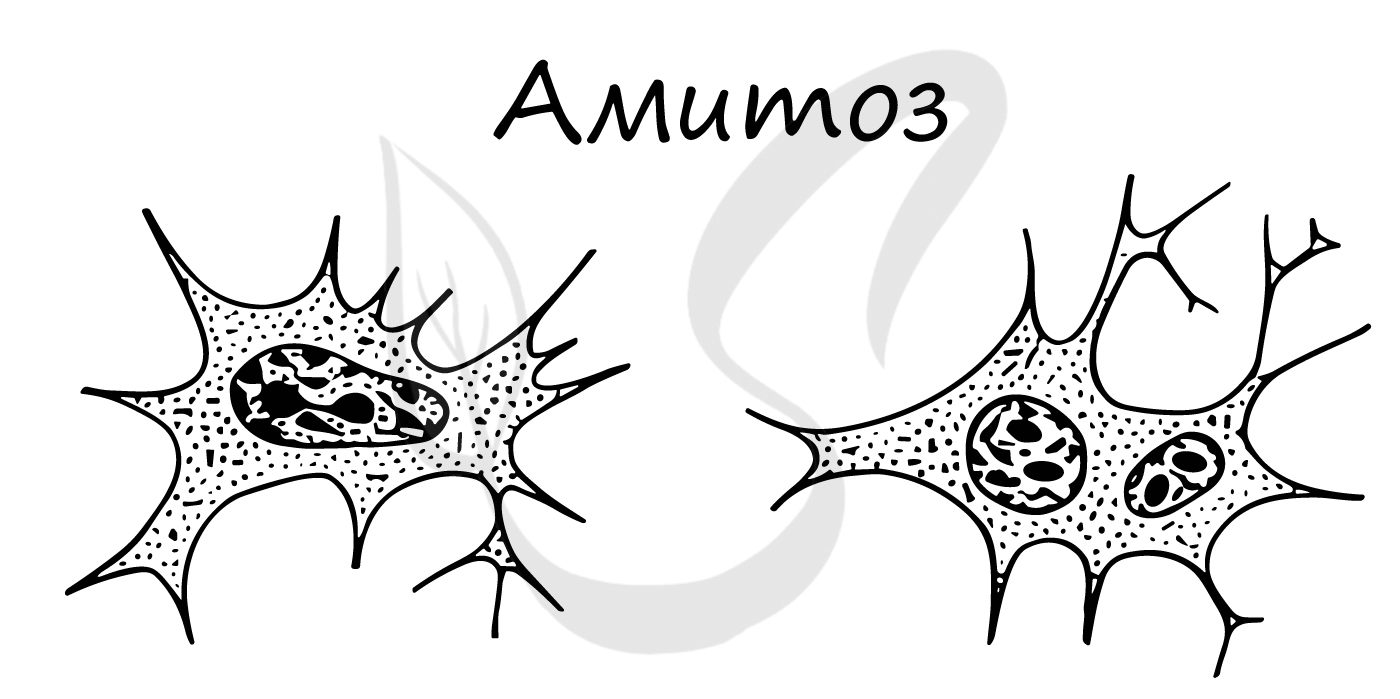
Амитоз встречается в раковых (опухолевых) клетках, воспалительно измененных, в старых клетках.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Что такое конъюгация хромосом
Каждая клетка в организме человека имеет двойной набор хромосом — один от отца и один от матери. Его обозначают «2N» и называют диплоидным. В сперматозоиде и яйцеклетке содержится одинарный набор хромосом, обозначаемый «1N» и называемый гаплоидным.
Процесс образования гаплоидного набора из диплоидного, происходящий при формировании половых клеток, называют мейозом. В пересчёте на количество центромер происходит сначала редукционное деление (мейоз I), а затем эквационное деление (мейоз II). У мужчин мейоз проходит так же, как и у большинства диплоидных видов, а у женщин данный процесс имеет некоторые отличия.
Кроссинговер между хромосомами отца и матери обеспечивает перегруппировку генетической информации между поколениями. Во время оплодотворения происходит слияние гаплоидных наборов хромосом сперматозоида и яйцеклетки, таким образом в зиготе восстанавливается диплоидный набор.
Мейоз I
Мейоз I имеет много общего с митозом, однако это более сложный и продолжительный процесс.
Мейоз первичных сперматоцитов и овоцитов начинается после фазы G2 митоза, а потому они имеют диплоидный набор хромосом (2N), содержащих реплицированную ДНК в составе сестринских хроматид (4С). Профаза I включает обоюдный обмен между хроматидами матери и отца при помощи крос-синговера.
Профаза I
• Лептотена. Хромосомы представлены в виде длинных нитей, прикреплённых концами к ядерной оболочке.
• Зиготена. Хромосомы сокращаются, образуют пары и гомологи слипаются друг с другом (синапсис). Данный процесс характеризует точное совмещение хромосом (ген к гену на протяжении всего генома). При этом у первичных сперматоцитов хромосомы X и Y образуют синапсис только концами своих коротких плечей.
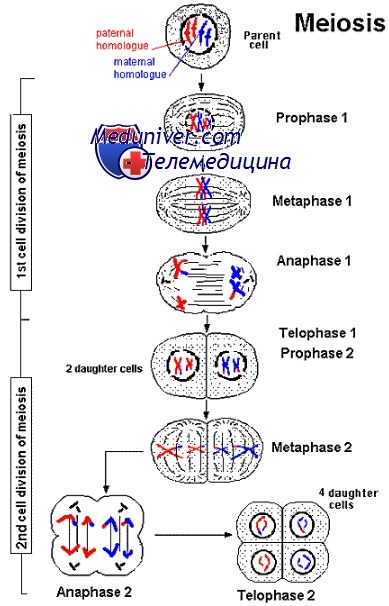
• Пахитена. Сестринские хроматиды начинают разделяться. Пары гомологичных хромосом, называемые бивалентами, имеют по четыре двойных спирали ДНК (тетрада). Одна или обе хроматиды каждой из отцовских хромосом скрещиваются с материнскими и образуют синаптонемальный комплекс. Каждая пара хромосом претерпевает хотя бы один кроссинговер.
• Диплотена. Происходит разделение хроматид, за исключением участков кроссинговера, или хиазм. Хромосомы всех первичных овоцитов находятся в таком состоянии вплоть до овуляции.
• Диакинез. Реорганизованные хромосомы начинают расходиться. В этот момент каждый бивалент содержит четыре хроматиды, соединённые обыкновенными центромерами, и несестринские хроматиды, соединённые хиазмами.
Метафаза I, анафаза 1, тепофаза 1, цитокинез I
Данные стадии мейоза подобны фазам митоза. Основное отличие: вместо разъединения несестринских хроматид происходит распределение по дочерним клеткам парных кроссоверных сестринских хроматид, соединённых центромерами.
В конце мейоза I вторичные сперматоциты и овоциты имеют 23 хромосомы (1N), каждая из которых состоит из двух хроматид (2С).
Мейоз II
При мейозе II возникает кратковременная интерфаза, во время которой не происходит репликации хромосом. Затем следуют профаза, метафаза, анафаза, телофаза и цитокинез. Схожесть каждой фазы мейоза II с подобной ей при митозе заключается в том, что пары хроматид (биваленты), соединённые в области центромер, выстраиваются в линию и образуют метафазную пластинку, а затем расходятся по дочерним клеткам, после чего следует репликация ДНК центромер.
В конце мейоза II в клетках содержится 23 хромосомы (IN), каждая из которых состоит из одной хроматиды (1С).
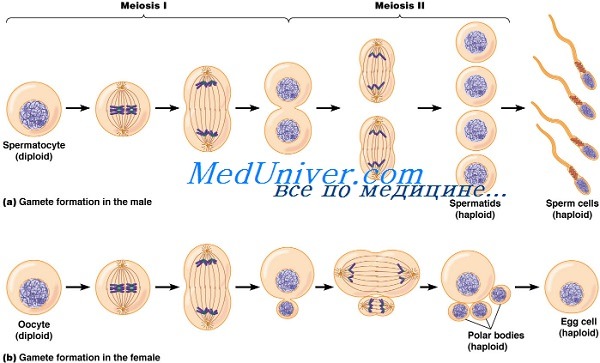
Мейоз у мужчин
Сперматогенезом называют процесс длительностью до 64 дней, включающий все стадии, на протяжении которых сперматогоний превращается в сперматозоид. При этом цитокинез остаётся незавершённым, позволяя каждому поколению клеток быть связанным цитоплазматическими мостиками.
После того как диплоидный первичный сперматоцит проходит стадию мейоза I, возникают два гаплоидных вторичных сперматоцита. Затем следует мейоз II, в результате которого появляются четыре гаплоидные спсрматиды. Во время спермиогенеза сперматиды превращаются в сперматозоиды. Данный процесс включает:
— образование акросомы, содержащей ферменты, которые способствуют проникновению семени;
— конденсацию ядра;
— удаление большей части цитоплазмы;
— формирование шейки, средней части и хвоста.
Мейоз у женщин
Овогенез начинается у плода в возрасте 12 недель и внезапно прекращается к 20-й неделе. Первичные овоциты остаются в форме диплотены профазы I вплоть до овуляции. Данную стадию называют диктиотеной.
Обычно созревает не более одного овоцита в месяц. Под влиянием гормонов первичный овоцит набухает, накапливая цитоплазматический материал. По завершению мейоза I он наследуется одной дочерней клеткой — вторичным овоцитом. Второе ядро переходит в первое направительное тельце, которое обычно не делится и со временем дегенерирует. После окончания мейоза I вторичный овоцит попадает в матку или фаллопиевы трубы.
Мейоз II вторичного овоцита останавливается на стадии метафазы до попадания в него сперматозоида. После этого процесс деления завершается, и образуется большой гаплоидный пронуклеус яйцеклетки, который сливается с пронуклеусом сперматозоида, а также второе направительное тельце, которое дегенерирует.
В зависимости от того, когда произойдёт оплодотворение, продолжительность данного процесса составляет 12—50 лет.
Медицинское значение понимания мейоза
• Диплоидный набор хромосом соматических клеток снижается до гаплоидного в половых клетках.
• Отцовские и материнские хромосомы подвергаются пересортировке, в результате чего количество возможных комбинаций (за исключением рекомбинаций внутри самих хромосом) возрастает до 223 (8 388 608).
• Пересортировка отцовских и материнских аллелей внутри хромосом создаёт между гаметами бесконечное количество генетических вариаций.
• Случайность процесса пересортировки отцовских и материнских аллелей во время мейоза (и оплодотворения) позволяет применять теорию вероятностей к генетическим пропорциям и генетической изменчивости согласно законам Менделя.
• Частота кроссинговера внутри хромосом позволяет предположить относительное расположение того или иного гена.
• Ошибки, возникающие во время конъюгации хромосом и кроссинговера, могут вызвать транслокации, разделение или расхождение хромосом, что часто становится причиной анеуплоидии.
Учебное видео: мейоз и его фазы

Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в содержание раздела «генетика» на нашем сайте
Биология. 11 класс
§ 18. Мейоз
Мейоз — особый способ деления эукариотических клеток, в результате которого из одной материнской клетки образуются четыре дочерние с уменьшенным в 2 раза набором хромосом. Если в мейоз вступает диплоидная клетка (2n4c), то образуются четыре гаплоидные клетки (1n1c). Клетки с гаплоидным набором хромосом не способны делиться мейозом.
Мейоз представляет собой два последовательных деления — мейоз I и мейоз II. Важно отметить, что репликация ДНК предшествует только первому мейотическому делению. Между мейозом I и мейозом II удвоения ДНК не происходит. Каждое из двух делений обычно включает профазу, метафазу, анафазу и телофазу. Рассмотрим процесс мейотического деления диплоидной клетки. Первое деление мейоза осуществляется следующим образом (табл. 18.1)
Таблица 18.1. Первое деление мейоза (мейоз I)
Схема
Фаза и процессы, происходящие в ней
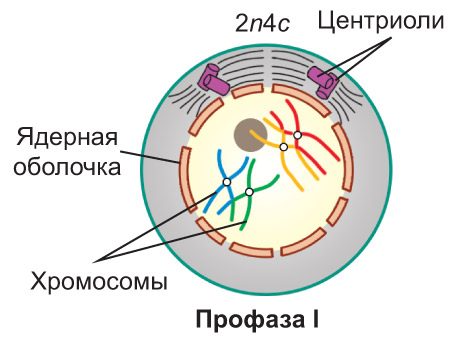
Гомологичные хромосомы попарно сближаются и соединяются друг с другом *сначала в нескольких участках, а затем по всей своей длине*. Этот процесс называется конъюгацией гомологичных хромосом, *а образовавшиеся хромосомные пары — бивалентами. В ходе конъюгации хромосомы тесно соприкасаются. В некоторых точках соприкосновения, называемых хиазмами, вследствие разрыва и последующего воссоединения молекул ДНК,* между хроматидами гомологичных хромосом может происходить обмен соответствующими участками — кроссингóвер. *Далее гомологичные хромосомы в составе каждого бивалента начинают отталкиваться друг от друга и в результате остаются связанными только в областях хиазм.*
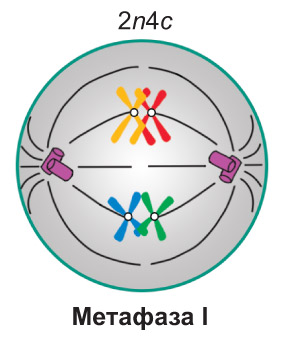
Метафаза I. Завершается формирование веретена деления. Спирализация хромосом достигает максимума. * Биваленты * располагаются в центральной части клетки, *образуя метафазную пластинку. Каждый бивалент ориентирован таким образом, что центромеры гомологичных хромосом находятся по разные стороны от экваториальной плоскости клетки, на одинаковом расстоянии от нее.* При этом нити веретена деления, идущие от противоположных полюсов клетки, прикреплены к центромерам разных гомологичных хромосом. Таким образом, в составе каждого *бивалента* одна из хромосом оказывается связанной с одним полюсом клетки, а другая — с противоположным
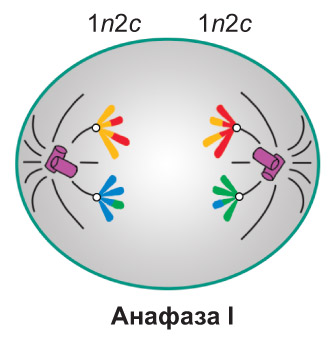
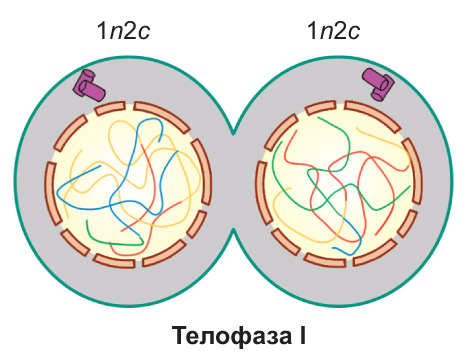
Телофаза I. Веретено деления разрушается. Происходит деспирализация хромосом и формирование двух ядер. Далее клетка разделяется на две дочерние. Они имеют гаплоидный набор хромосом, каждая хромосома состоит из двух хроматид (1n2c)
Промежуток времени между первым и вторым делениями мейоза обычно очень короткий. В этот период, как уже отмечалось, не осуществляется репликация ДНК. Каждая из двух клеток, образовавшихся в результате мейоза I, вступает в мейоз II. Это деление протекает аналогично митозу (табл. 18.2).
*У некоторых организмов перерыв между мейозом I и мейозом II — так называемый интеркинез вообще отсутствует. При этом клетки, завершившие телофазу I, сразу же переходят в профазу II. Более того, могут выпадать и эти фазы. Так, у большинства растений, клетки, делящиеся мейозом, после окончания анафазы I вступают непосредственно в метафазу II. В этом случае переход от первого мейотического деления ко второму не сопровождается деспирализацией хромосом, формированием двух ядер и разделением материнской клетки на две дочерние.*
Таблица 18.2. Второе деление мейоза (мейоз II)
Схема
Фаза и процессы, происходящие в ней
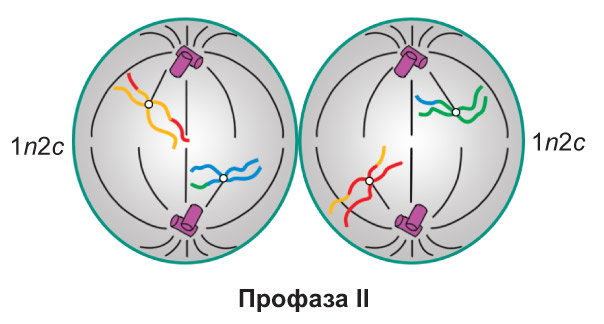
Профаза II. В результате спирализации хроматина формируются двухроматидные хромосомы (набор 1n2c). В это же время начинается образование веретена деления. После распада ядерной оболочки отдельные хромосомы беспорядочно располагаются в гиалоплазме
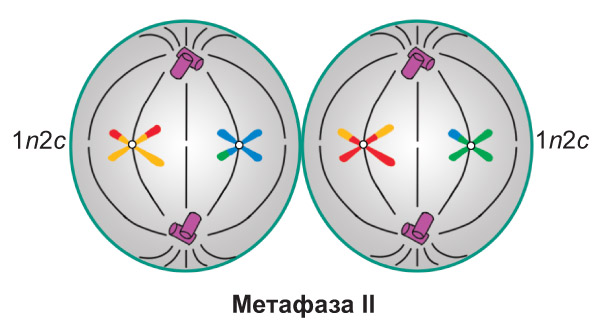
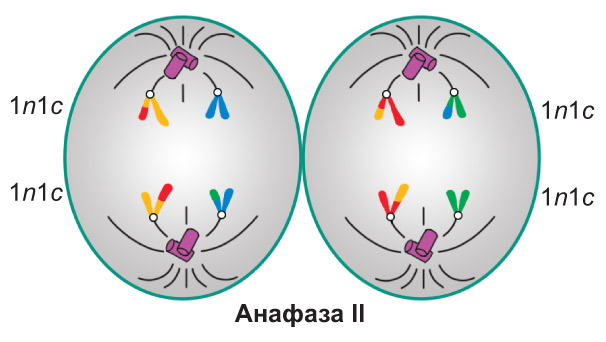
Анафаза II. Происходит разделение центромер. Сестринские хроматиды (теперь уже дочерние хромосомы ) растягиваются к разным полюсам клетки. В конце анафазы II набор хромосом и хроматид у каждого полюса составляет 1n1c
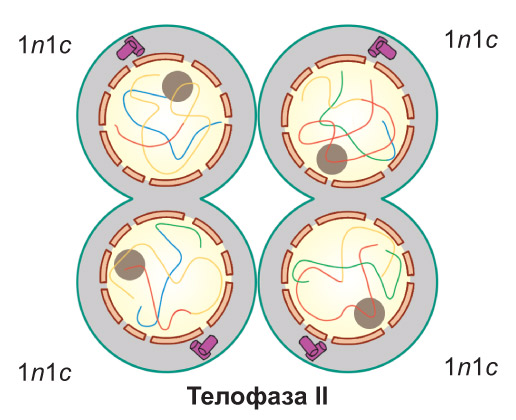
Телофаза II. Хромосомы деспирализуются, формируются ядра, и происходит разделение клеток. При этом образуются четыре дочерние клетки, имеющие набор 1n1c
Таким образом, в результате первого деления мейоза исходная материнская клетка (2n4c) разделилась на две дочерние, имеющие гаплоидный набор хромосом. *Следовательно, в мейозе I произошла редукция (уменьшение) числа хромосом в 2 раза. Поэтому первое деление называют редукционным. В образовавшихся гаплоидных клетках каждая хромосома представлена двумя хроматидами (1n2c).
В результате второго деления, протекающего по типу митоза, набор хромосом (1n) не изменяется. Это деление мейоза называется эквационным (от лат. aequatio — уравнивание). Эквационное деление, в отличие от редукционного, сопровождается расхождением сестринских хроматид. Поэтому итогом второго деления является образование четырех гаплоидных клеток с однохроматидными хромосомами (1n1c).*
*Как и в случае митоза, правильное протекание мейоза может быть нарушено действием определенных внешних или внутриклеточных факторов. Последствия таких воздействий могут быть разными, вплоть до образования нежизнеспособных дочерних клеток. Наиболее часто встречающейся патологией мейоза является нерасхождение хромосом в анафазе I. Оно происходит вследствие нарушения разделения бивалентов и выражается в том, что обе гомологичные хромосомы перемещаются к одному из полюсов клетки. Нерасхождение может наблюдаться и на стадии анафазы II. При этом к одному и тому же полюсу отходят две сестринские хроматиды (дочерние хромосомы ). Как в первом, так и во втором случае результатом нерасхождения хромосом является образование генетически неравноценных клеток. В одних клетках наблюдается избыток хромосом (1n + 1), а в других — недостаток (1n — 1).*
Биологическое значение мейоза. У животных и человека путем мейоза образуются гаметы — гаплоидные половые клетки. В результате последу ющего оплодотворения формируется зигота с двойным набором хромосом, из которой развивается новый организм. Он является диплоидным, как и его родители, а значит, сохраняет свойственный данному виду организмов кариотип. Без мейоза, приводящего к уменьшению набора хромосом в 2 раза, половое размножение сопровождалось бы удвоением числа хромосом в каждом новом поколении. У растений, многих водорослей и грибов мейоз приводит к формированию спор, с помощью которых осуществляется бесполое размножение.



