Клада
В русскоязычной литературе встречается также другой вариант транслитерации этого слова — «клад».
Любая клада рассматривается как монофилетическая группа организмов и может быть изображена с помощью кладограммы (диаграммы происхождения организмов в форме дерева, «родословной»).
На начальном этапе кладистического анализа утверждение о том, что данная группа организмов представляет собой кладу, выдвигается первоначально как научная гипотеза об эволюционных взаимоотношениях между организмами, участвующими в анализе. Данная гипотеза может быть поддержана или опровергнута последующим анализом, использующим другой набор данных.
Если гипотеза о монофилии группы оказывается поддержанной в различных кладистических исследованиях, использующих разные наборы данных, данной группе может быть придан статус таксона. Не все таксоны, которые используются в настоящий момент, соответствуют кладам (пресмыкающиеся, например, — парафилетическая группа, потому что она не включает птиц, которые тоже эволюционировали от общего предка рептилий). Однако в кладистике — в отличие от эволюционной таксономии — одной из приоритетных целей провозглашается такая реорганизация таксонов, чтобы они по возможности точно соответствовали кладам.
Связанные понятия
Упоминания в литературе
Связанные понятия (продолжение)
Синапсѝды (лат. Synapsida), также тероморфы или зверообразные (лат. Theromorpha, от др.-греч. θήρ, θηρίον — зверь + μορφή — форма) — таксономическая группа в составе клады амниот, возникшая около 318 млн лет назад. Одна из групп синапсид — цинодонты (Cynodontia) — была предком млекопитающих.
Кладистика

Кладистика (от др.-греч. κλάδος (kládos) — ветвь) — направление филогенетической систематики. Характерные особенности кладистической практики состоят в использовании так называемого кладистического анализа (строгой схемы аргументации при реконструкции родственных отношений между таксонами), строгом понимании монофилии и требовании взаимно-однозначного соответствия между реконструированной филогенией и иерархической классификацией. Кладистический анализ — основа большинства принятых в настоящее время биологических классификаций, построенных с учётом родственных отношений между живыми организмами.
Кладистике противостоят фенетика, основанная на количественной оценке так называемого общего сходства (англ. overall similarity ), и так называемая эволюционная таксономия, признающая ценность выяснения родственных отношений, но не требующая строгого соответствия системы и филогении (в частности, это выражается в признании права на существование в системе парафилетических групп).
Кладистика основана на идеях немецкого энтомолога и систематика Вилли Хеннига (1913—1976), изложенных в его работах 1950—1960-х гг., однако название кладистика было впервые использовано только в конце 1960-х годов критиками этого направления филогенетической систематики, а современные формализованные процедуры были разработаны только в 1970-е гг. его последователями в США.
Содержание
Определение


Характеристический признак, который есть как у корневого вида, так и у его потомков, называется плезиоморфия («близкая форма», также называется «предшествующей формой»). Характеристический признак, существующий только у позднейших потомков, называется «апоморфия» («обособленная форма», также называется «производной формой») для этой группы. Прилагательные «плезиоморфный» и «апоморфный» используются вместо «примитивный» и «передовой» (или «продвинутый») для предотвращения оценочного суждения об эволюционных формах, поскольку оба они могут обозначать структуры, крайне полезные при некоторых обстоятельствах и вредными — при других. Нередко набор плезиоморфных форм неформально называется «базисом» для клады или клад, которым этот набор присущ.
Несколько других распространенных терминов кладистики введены для описания кладограмм и позиций таксонов внутри них. Вид или клада является базальным относительно другой клады, если первая имеет больше плезиоморфных черт, чем вторая. Базальная группа насчитывает меньшее количество видов по сравнению с развитыми группами; наличие в кладограмме базальной группы не является обязательным. Например, при совместной кладистичной классификации птиц и млекопитающих, одна из этих групп не является базальной для другой.
Клада или вид, находящийся в кладограмме внутри другой клады, может называться «вложенной» или «вложенным» в эту кладу.
История названия
Классификационный подход, который сейчас известен как кладистика, был заложен работами Вилли (Вильгельма) Хеннига. Но его главная книга, даже в редакции 1979 года, не содержит собственно термина «кладистика». В этой книге автор ссылается на свой собственный подход как на «филогенетическую систематику», имея в виду название книги (Хенниг, 1979). Обзорная статья Дупиуса (1984) утверждает, что термин «клада» был предложен в 1958 году Джулианом Хаксли, «кладистика» Кейном и Харрисом в 1960-м, и «кладист» (для обозначения последователя школы Хеннига) Майером в 1965 году. Некоторые дебаты, куда были привлечены кладисты, продолжались с 19-го века, но кладисты присоединились к ним позже и с новыми взглядами, что описано в предисловии к книге Хеннига редакции 1979-го года, где Розен, Нельсон и Патерсон отмечали следующее:
 | Отягощенный неясными и скользкими идеями об адаптации, эволюционной пригодности, биологических видах и естественном отборе, неодарвинизм (который определяется как «эволюционный» подход к биологической систематике Майером и Симпсоном) не имеет определенного метода исследования, а также пришел в зависимость от консенсуса авторитетов по вопросам эволюционной интерпретации и классификации. |  |
Методы кладистики

Массив информации, который может быть подвергнут кладистическому анализу, должен быть организован специальным образом. Для этого, прежде всего, необходимо провести разграничение между признаками (или характеристикам) и их состояниями (характеристическими состояниями). Например, цвет перьев может быть голубым у одного вида и красным у другого. Тогда «голубые перья» и «красные перья» будут двумя состояниями одного признака — «цвета перьев».
Исследователь должен определить, какой признак (или признаки) присутствовал до появления последнего общего предшественника (плезиоморфия), а какой имелся у последнего общего предшественника (синапоморфия) путем выделения одного или нескольких корневых видов. Корневой вид — это организм, который не относится к исследуемой группе, но является ему близкородственным. Это делает выбор корневого вида важной задачей, поскольку такой выбор способен серьезно повлиять на структуру кладистического дерева. Надо заметить, что при характеристике клад используются только синапоморфии.
Применение гомоплазии в морфологических наборах данных часто можно избежать путем точнейшего определения характерных признаков и увеличения их количества: в предыдущем примере, используя в качестве признаков «крылья с перьями», «крылья с хитиновым экзоскелетом» и «кожистые крылья» как характерные признаки, можно избежать эволюционно ложного объединения трех перечисленных групп животных на основе гомоплазии. При анализе «супердеревьев» (баз данных, включающих большее число таксонов исследуемой клады), применение неточных признаков может стать неизбежным, поскольку в обратном случае признаки могут стать неприменимыми для всех многочисленных таксонов. Скажем, такой признак, как наличие крыльев, не может быть применен для анализа филогении настоящих многоклеточных животных (Eumetazoa), так как большинству видов этого таксона данный признак не присущ. Таким образом, осторожный выбор и определение характерных признаков является другим важным элементом кладистического анализа. При ошибочном определении корневого вида и набора признаков, никакие методы построения кладограмм не смогут дать в результате филогенетической системы, которая соответствует эволюционной реальности.
Для практически любого таксона можно построить не одну, а множество различных кладограмм, основываясь на различных корневых видах и наборах характерных признаков; но из них выбирают одну-единственную, руководствуясь принципом парсимонии : компактную систему, которая, в сочетании с наименьшими возможными изменениями характерных признаков (синаптоморф), дает непротиворечивую картину происхождения клады (в общем, это вариант соображения по бритве Оккама). Хотя в начале такой анализ проводился «вручную», впоследствии для него стали применяться компьютеры со специальным программным обеспечением, которое позволяет оперировать на порядки большими наборами данных и количеством признаков. Такие программы (вроде PAUP и других подобных) позволяют делать статистическую оценку вероятности нод (узлов и разветвлений) построенной кладограммы.
Важно также заметить, что ноды кладограммы не обязательно отражают различия эволюционных ветвей, а лишь различия постоянных признаков, которые наблюдаются между этими ветвями. Признаки, которые заключаются в разнице последовательностей ДНК, способны расходиться после того, как генный дрейф между популяциями редуцируется к некоторой пороговой величине, в то время как заметные морфологические изменения, обычно будучи эпистатическими (то есть результатом взаимодействия нескольких генов) выявляются только после того, как разошедшиеся таксоны отдельно эволюционно развивались в течение некоторого (обычно довольно значительного) времени; так, биологические подвиды зачастую могут быть разрознены генетически, но не морфологически (по строению тела или внутренней анатомии).
В идеале морфологические, молекулярно-биологические и другие (поведенческие, экологические, палеонтологические, и т. д.) филогенетические данные должны обобщаться при разработке итогового заключения, при этом никакой из методов не является более доказательным, чем другие, но все они имеют разные внутренние источники ошибок. Например, конвергенция развития признаков (гомоплазия) гораздо чаще появляется при анализе морфологических данных, чем в данных, полученных при молекулярном секвенировании, но реверсии признаков у них обоих встречаются с примерно одинаковой частотой; обычно, морфологические гомоплазии могут быть вскрыты при достаточно внимательном и детальном анализе характерных признаков.
Кладистика не признает ни одну из имеющихся теорий эволюции, принимая во внимание только общую информацию о происхождении с существенными ее модификациями. Благодаря этому кладистические методы могут быть (и бывают) использованными в небиологических дисциплинах, включая историческую лингвистику и задачи по определению авторства текстов.
Кладистическая классификация

Узловой: недавний совместный предок видов А и Б и все его потомки
Ветвяной: все потомки древнейшего общего предка А и Б, которые одновременно не являются предками В
Апоморфный: древнейший общий предок А и Б, у которого имеется некоторая определяющая апоморфия, и все его потомки
С 1960-х в биологии возникла тенденция под названием «кладизм» или «кладистическая таксономия» считать таксоны кладами. Иными словами, кладисты считают, что биологическая классификация должна быть реформирована таким образом, чтобы устранить все не-клады. В отличие от кладистов, все остальные таксономисты настаивают на том, что каждая из таксономических групп живых существ отражает филогенетическую историю. Последние часто используют кладистический подход, но также допускают в формировании классификационных деревьев использование как монофилетических (что совпадает с кладистичним подходом), так и парафилетических таксонов. Как результат, уже как минимум с начала XX века роды и таксоны низшего уровня формировали основываясь на монофилетическом подходе, в то время как таксоны высокого ранга могут быть (а такие как класс и выше — обычно и являются) парафилетическими.
В данной системе упорядочения монофилетической группой считается клада, состоящая из предшественника и всех его потомков, которые формируют одну и только одну эволюционную группу. Парафилетической является такая же, но без некоторых потомков, которые претерпели существенные изменения. Например, традиционный класс Пресмыкающиеся не включает птиц, хотя птицы произошли от пресмыкающихся; аналогично, традиционная группа Беспозвоночные является парафилетической, потому что не включает позвоночных, которые происходят от неё.
Группа, состоящая из членов, которые происходят из разных эволюционных линий, называется полифилетической. Например, сформированная ранее таксономическая группа «толстокожие» (Pachydermata) была затем признана полифилетической, поскольку включенные в нее слоны, носороги и бегемоты произошли от разных, не родственных между собой предшественников. Аналогично, сформированный прежде тип Кишечнополостные был позднее разделенн на книдарий и гребневиков на основании их происхождения от различных предшественников и возникших вследствие этого существенных морфологических различий. Эволюционные таксономисты считают полифилетические группы ошибками классификации, вызванными конвергенцией и другими видами гомоплазии, которые ошибочно интерпретируются как гомологии.
Следуя Хеннигу, кладисты считают, что парафилия является не менее неприемлемой для классификации, чем полифилия. В рамках кладистики считается, что монофилетические группы могут быть объективно и строго определенными, основываясь на общих предшественниках или синаптоморфах. В отличие от них, парафилетические и полифилетические группы выделяются на основе анализа ключевых характеристик, в ходе которого степень важности этих характеристик является весьма субъективной. Кладисты отмечают, что, таким образом, это приводит к «градистическому» мышлению, которое в свою очередь тяготеет к телеологии, которую обычно считают неприемлемой в эволюционных исследованиях, поскольку телеология подразумевает существование общего плана эволюции, который не может быть показан экспериментально.
Продолжая эту линию критики, некоторые кладисты утверждают, что таксоны высшего по виду ранга являются слишком субъективными, и поэтому их выделение не представляет никакой содержательной информации. На основе этого, как утверждают такие критики, эти таксоны не должны использоваться в систематике вообще. Таким образом, биологическая систематика, в их понимании, должна полностью отойти от Линнеевской таксономии и стать простой иерархией клад. Весомость этой системы аргументов критически зависит от того, как часто в ходе эволюции прерывается градуалистическое равновесие экологических условий. Квази-стабильное состояние экосистем приводит к медленному филогенезу, который может быть трудно интерпретирован в рамках линнеевской систематики; в то же время катастрофическое событие выводит таксон из экологического равновесия и обычно приводит к разделению признаков клад. Это разделение происходит в течение эволюционно короткого времени, и в результате дает систему таксонов, которая легко описывается в рамках линнеевской классификации.
Другие эволюционные систематики утверждают, что все таксоны по своей сути субъективны, даже если они отражают эволюционные взаимоотношения, поскольку живые организмы формируют непрерывное эволюционное дерево. В таком случае, любая линия раздела является искусственной и создает монофилетическое выделение над парафилетическим. Парафилетические таксоны являются необходимыми для классификации более ранних секций дерева — например, ранние позвоночные, которые через некоторое время развились в семью Гоминиды, не могут быть помещены более в одну монофилетическую семью. Также в этой системе аргументов приводится то, что парафилетические таксоны предоставляют информацию о существенных изменениях в морфологии, экологии и эволюционной истории организмов — короче говоря, что и таксоны, и клады являются ценными для построения истинной картины систематики живой природы, но и те, и те — с некоторыми ограничениями.
В полемике вокруг годности кладистических методов при построении биологической классификации термин «монофилия» часто используется в своем старом смысле, где он включает в себя парафилию, наряду с чем используется термин «голофилия» для описания «чистых» клад («монофилии» в Хеннингском смысле). Как пример ненаучности эмпирического метода используется следующий: если какая-то парафилетическая эволюционная линия приводит к возникновению клады из-за значительной адаптивной радиации и многочисленных, хорошо заметных и важных синапоморфий, особенно таких, что ранее в этой группе живых существ не встречались, парафилия не признается достаточным аргументом против выделения таксона в линнеевской системе классификации (но это считается достаточным контраргументом в рамках филогенетической номенклатуры). Например, как было указано выше, и завропсиды («рептилии»), и птицы в линнеевской систематике имеют одинаковый ранг классов, хотя последние являются эволюционно крайне специализированной ветвью рептилий.
Формальный сборник филогенетической номенклатуры (ФилоКод) сейчас находится в разработке для приспособления его к кладистической таксономии. Его планируют использовать и те, кто пытается полностью избегать линнеевской номенклатуры, и те, кто использует в систематике таксоны вместе с кладами. При некоторых обстоятельствах (например, при классификации гесперорнисов) методы ФилоКода были привлечены во избежание неопределенностей, которые возникали при применении линнеевской систематики к этому таксону; таким образом, комбинация этих двух систем позволяет сформировать таксономическую картину, которая размещает группу живых существ на эволюционном дереве в полном соответствии с имеющейся научной информацией.
Клада человека и её первые представители
Клада человека и её первые представители
Если оценивать значимость эволюционных процессов с антропоцентристской точки зрения, то наиболее важным событием в позднемиоценовой истории гоминоидов было, конечно, появление первых представителей семейства гоминид (Hominidae), и отделение гоминидной линии эволюции, ведущей к человеку, от общего с другими человекообразными обезьянами генеалогического ствола. Люди и те их предки (как прямые, так и находящиеся на боковых или тупиковых ветвях нашего генеалогического древа), которые жили уже после того, как совершилось это филогенетическое расхождение, составляют группу, часто обозначаемую как клада человека (human clade — англ.). Она включает несколько родов и примерно два десятка видов. Преобладающие в настоящее время (на конец 2002 г.) представления о её составе, а также эволюционной истории суммированы в таблице 1 и на рисунках 9 и 10.
Таблица 1. Таксономический состав клады человека, или семейства гоминид, согласно представлениям большинства антропологов на середину 2003 г.
Рис. 9. Клада шимпанзе (верхняя ветвь) и клада человека (нижняя ветвь)
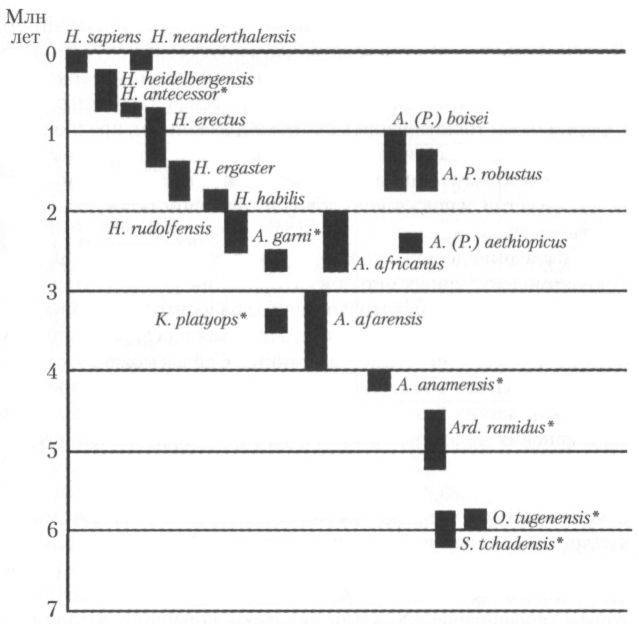
Рис. 10. Время существования разных видов гоминид. Степень филогенетической близости к Homo sapiens убывает слева направо. Звёздочками отмечены таксоны, выделенные после 1994 г.
Следует подчеркнуть, что термин гоминиды здесь и далее употребляется именно и только для обозначения членов клады человека. Ещё недавно такое его понимание было почти общепринятым и не нуждалось в оговорках. Однако в последние годы, вследствие развития биомолекулярных методов реконструкции филогенеза, с одной стороны, и внедрения в биологическую систематику ряда новых методов и правил, с другой, в семейство гоминид часто стали включать ещё и африканских человекообразных обезьян, а иногда, в качестве подсемейства, и орангутанга и даже гиббона. Таксономический ранг группы, включающей только членов клады человека, при этом, соответственно, понижается. Иногда её выделяют как подсемейство с названием Homininae, иногда как трибу Hominini, а иногда как род Homo, или даже подрод Homo homo. Тем не менее многие антропологи и палеонтологи, считая такой пересмотр классификационных схем с формальной точки зрения в общем оправданным, предпочитают всё же, во избежание путаницы, применять термин «гоминиды» по-прежнему лишь по отношению к представителям человеческой ветви, и пока такое словоупотребление остаётся наиболее распространённым.
В последние десять лет, благодаря, с одной стороны, развитию биомолекулярных методов датирования филогенетических событий, а с другой, некоторым новым палеонтологическим находкам, удалось существенно продвинуться в решении вопроса о времени рождения семейства гоминид. Сейчас можно уже почти не сомневаться в том, что эволюционные пути наших предков и предков наших ближайших родственников — современных африканских человекообразных обезьян, — разошлись где-то в интервале от 8 до 4,5 млн лет назад.
Первая попытка определить время расхождения эволюционных линий гоминид и других человекообразных обезьян молекулярными методами была предпринята А. Уилсоном и В. Сэйришем из университета Калифорнии в Беркли (США). В 1967 г. они опубликовали результаты своих исследований, согласно которым генеалогические ветви человека, шимпанзе и гориллы разделились где-то в интервале от 4 до 6 млн лет назад. В 1987 г. Ч. Сибли и Дж. Алквист из другого американского университета — Йельского, привели данные, свидетельствующие о том, что горилла отделилась от общего ствола раньше, чем шимпанзе, и постепенно накапливающиеся новые молекулярные и палеонтологические данные заставляют большинство специалистов думать, что, скорее всего, так оно и было (табл. 2). Впрочем, для полноты картины следует упомянуть ещё весьма экзотическую и мало кем поддерживаемую гипотезу, согласно которой шимпанзе и горилла являются потомками ранних гоминид — австралопитеков, оставившими прямохождение и вторично приспособившимися к передвижению на четырёх ногах.
Таблица 2. Оценки времени расхождения эволюционных линий человекообразных обезьян и человека, даваемые разными исследователями на основании молекулярных данных. Для сравнения, там, где возможно, дана оценка по палеонтологическим данным. (Все даты в млн лет)
Орангутанг Горилла Шимпанзе Sarich, Wilson 1967 5 Sibley, Alquist 1987 7,7–11 5,5–7,7 Holmes et al. 1988 16,2–9,2 5,9–8,9 Hasegawa et al. 1990 9–15 4,3–5,9 3,2–4,6 Adachi, Hasegawa 1995 5,1–6,5 4–5 Estreal et al. 7,3–9,8 5 3,2–4,5 Horai et al. 1992 7,0–8,4 4,2–5,2 Horai et al. 1995 6,3–6,9 4,7–5,1 Ruvolo 1995 7,4–8,9 5–6 Hedges et al. 2001 4,5–6,5 По палеонтологическим данным 15 >4,5
Что касается генеалогических корней семейства гоминид, то их поиск ещё далёк от завершения. Череда сенсационных открытий, начавшихся в 1994 г. и продолжающихся по сей день, сильно усложнила и даже запутала уже, казалось бы, прояснявшуюся в общих чертах картину. С одной стороны, эти открытия подтвердили высказывавшееся некоторыми исследователями предположение, что род австралопитеков (о нём подробней говорится в следующем разделе), представляющий собой одно из важнейших звеньев, связывающих человекообразных обезьян с человеком, не является всё же древнейшим звеном. С другой стороны, о его предшественниках на этом пути известно до сих пор очень немного. Найти и, главное, идентифицировать останки самых первых представителей клады человека оказалось сложнее, чем определить примерное время их появления. В какой-то степени это объясняется тем, что в Африке местонахождения ископаемой фауны, относящиеся к тому периоду, когда должно было совершиться выделение гоминидной линии эволюции (поздний миоцен — ранний плиоцен), довольно редки. Кроме того, недостаток материалов усугубляют и методические трудности их анализа. Даже если допустить, что скелетные останки членов предковой для всего семейства гоминид группы уже обнаружены, точно определить их, выделить среди других похожих находок — зубов, обломков челюстей, мелких фрагментов костей черепа или конечностей — всё равно было бы очень трудно.
До конца 70-х гг. XX в. считалось, что вполне приемлемым и наиболее перспективным претендентом на весьма почётное место в самом основании генеалогического древа гоминид является известный по находкам в Южной Азии и в Европе род рамапитек (Ramapithecus). В таковом качестве он до сих пор иногда фигурирует в отечественной научно-популярной и даже учебной литературе. Однако за последние два десятилетия в результате появления новых материалов и переоценки старых, филогенетическая роль рамапитека подверглась решительному пересмотру. Большинство специалистов теперь считает, что рамапитек, живший в период примерно от 15 до 7 млн лет назад, не имеет непосредственного отношения к происхождению гоминид. Он стоит, скорее, у истоков линии, ведущей к орангутангу, а не к человеку, либо же вообще не связан прямо ни с тем, ни с другим.
Хотя вплоть до полевых открытий, сделанных в последние 3–4 года, о гоминоидах конца миоцена и начала плиоцена почти ничего не было известно, на роль, освободившуюся с «изгнанием» рамапитека, выдвигались и другие кандидаты. Наибольшую известность среди них получил упоминавшийся уже в предыдущей главе уранопитек (Ouranopithecus), иногда именуемый ещё грекопитеком (Graecopithecus). Фрагменты черепа и несколько зубов этого существа были обнаружены на севере Греции в отложениях возрастом около 10 млн лет. Некоторые исследователи видели в уранопитеке возможного родоначальника гоминид, предполагая, что эта форма могла быть распространена не только в Европе, но и в Африке, а её отсутствие среди африканских ископаемых гоминоидов объясняется просто плохой сохранностью материалов соответствующего периода. Однако признаки, которые сближают уранопитека и ранних гоминид — это примитивные черты, доставшиеся и тем и другим в наследство от их общих предков. Такие признаки не могут служить в качестве аргумента в пользу прямой связи между двумя данными группами.
Место в основании эволюционной линии гоминид и до сих пор остаётся вакантным, но конкуренция за то, чтобы его занять, на рубеже второго и третьего тысячелетий резко возросла. Это связано прежде всего с интенсификацией полевых исследований, проводимых в тех районах, где наиболее вероятно обнаружение скелетных останков древнейших предков человека. Американскими, английскими и особенно французскими экспедициями, постоянно работающими сейчас в Восточной и Центральной Африке, был сделан ряд открытий, не только углубивших на несколько миллионов лет нашу родословную, но и значительно расширивших круг претендентов на роль основателя клады человека.
Сначала очень интересные находки появились на севере Эфиопии, в давно уже облюбованном палеоантропологами и археологами регионе, известном под названием Средний Аваш. В 1994 г. были описаны остатки скелета, найденного на местонахождении Арамис, в геологическом слое, сформировавшемся около 4,4 млн лет назад. Этот материал находился к моменту открытия в очень плохом состоянии, и до сих пор были детально изучены только зубы и фрагменты черепа, а также головка плечевой и обломок лучевой костей. Анализ этих находок привёл сначала к провозглашению нового — самого древнего — вида из рода австралопитеков, но через несколько месяцев авторы первого описания костей из Арамис сочли их обладателя достойным более высокого — родового ранга и опубликовали поправку, в которой вместо вида Australopithecus ramidus представили миру род и вид Ардипитек рамидус (Ardipithecus ramidus).
Открытие рамидуса получило широкую известность, и многие антропологи рассматривают его как вполне достоверного гоминида. Однако иногда высказываются и сомнения относительно статуса этого существа. Некоторые специалисты считают проблематичным отнесение костей из Арамис к гоминидам, указывая на наличие ряда черт, сближающих ардипитека рамидус с шимпанзе (например, относительно крупные клыки). Тем не менее большинство признаков говорит всё-таки в пользу его принадлежности к гоминидам. Особенно важно, что затылочное отверстие у рамидуса находится близко к середине основания черепа, а это особенность, свойственная двуногим, т. е. прямоходящим существам. В какой-то мере ситуацию могут прояснить новые палеоантропологические находки, сделанные в Среднем Аваше в конце 90-х гг. Они включают обломок челюсти с зубами и фрагменты нижних и верхних конечностей, возраст которых составляет от 5,2 до 5,8 млн лет. Эти кости были описаны как подвид рамидуса (Ardipithecus ramidus kadabba).[10] Хотя и в данном случае нельзя пока исключать, что в лице нового подвида мы имеем дело не с ранним гоминидом, а с прародителем шимпанзе или общим предком и тех и других, принадлежность к гоминидам кажется наиболее вероятной. Дело в том, что, судя по строению кости одного из пальцев ноги, Ardipithecus ramidus kadabba был уже прямоходящим существом. Как останки рамидуса рассматривают иногда и обломок нижней челюсти, происходящей с местонахождения Табарин в Кении. Возраст этой находки составляет 4,4–4,5 млн лет.
Хотя на протяжении шести лет с момента открытия рамидус оставался древнейшим из известных антропологам членов клады человека, большинству исследователей было ясно, что с его обнаружением сюрпризы ещё далеко не закончились. Всего три года назад автор этих строк писал, что «материалы из Арамис, заполняющие последний, пожалуй, из остававшихся в палеонтологической “летописи” крупных разрывов между обезьяной и человеком, позволяют предполагать, что австралопитеки не были первыми гоминидами, и что рано или поздно могут быть обнаружены ещё какие-то промежуточные формы, сосуществовавшие с рамидусом, или даже предшествовавшие ему».[11] После ряда замечательных палеоантропологических находок, сделанных в 2000–02 гг., в справедливости этого предположения уже не приходится сомневаться.
Сначала, в 2000 г., очень древние кости, похожие на останки гоминида, были выявлены на местонахождениях Капсоним, Капчеберек и Арагай в местности Туген Хиллс на западе Кении. Зубы, фаланги и фрагменты бедренной и плечевой костей, обнаруженные здесь в слоях геологической формации Лукейно и датированные временем около 6 млн лет назад, позволили изучавшим их французским антропологам выделить новый род и вид вероятных дальних предков человека. Он получил название оррорин тугененсис (Orrorin tugenensis), образованное от имени низкогорья (Туген), где велись поисковые работы и от слова «оррорин», означающего на языке одного из местных племён «первый человек».[12] Чуть позже, в 2001–02 гг., на местонахождении ископаемой фауны Торос-Меналла в центральноафриканской пустыне Джураб другой французской экспедицией были найдены останки ещё одного гоминидоподобного существа, также, возможно, жившего примерно 6 млн лет назад, или даже несколько раньше. Среди его костей особенно важна хорошо сохранившаяся верхняя часть черепа (мозговая коробка плюс лицевой скелет), дополняемая обломком нижней челюсти с несколькими зубами. Этой форме по историческому названию той части Африки (Сахель) и той страны (Чад), откуда происходит материал, дали имя сахельантроп (Sahelanthropus tchadensis).[13]
Благодаря открытию ардипитека (Ardipithecus ramidus), oppoрина (Orrorin tugenensis) и сахельантропа (Sahelathropus tchadensis), стало окончательно ясно, во-первых, что гоминидная линия эволюции выделилась не менее 5 млн лет назад, и, во-вторых, что те несколько видов австралопитеков, которые ещё недавно считались нашими древнейшими предками, на самом деле далеко не исчерпывают собой всего разнообразия форм ранних гоминид. Вполне можно ожидать, что в скором времени количество вероятных позднемиоценовых и раннеплиоценовых членов клады человека возрастёт ещё более. Похоже, что уже с самого начала истории семейства гоминид оно было весьма многочисленным, но подавляющее большинство составлявших его родов и видов просуществовало сравнительно недолго, окончив свой путь в тупиках эволюционного лабиринта.
Как это очень часто бывает в науке, новые данные, проясняя одни вопросы, одновременно усложняют решение других. Особенно серьёзные проблемы возникают в результате выделения рода и вида оррорин (Orrorin tugenensis). Некоторые анатомические особенности этого таксона таковы, что, если признавать его реальность и принадлежность к гоминидам, то это может иметь следствием переоценку эволюционной роли рода австралопитеков вплоть до исключения их из числа предков рода Homo. Дело в том, что коренные зубы представителей вида оррорин, подобно зубам всех представителей рода Homo, к которому принадлежат современные люди, невелики по размеру (относительно реконструированной величины тела) и имеют толстую эмаль. У австралопитеков зубы также с толстой эмалью, но вот их относительный размер гораздо больше. Следовательно, по этому очень важному признаку (он важен, прежде всего, потому, что особенности строения зубов отражают особенности питания и поведения в целом) австралопитеки уклоняются от линии, связывающей древнейших и поздних гоминид, и при желании могут быть представлены как боковая ветвь на нашем генеалогическом древе.
Придётся ли в действительности предпринимать столь решительную перекройку существующих филогенетических схем, сейчас ещё далеко не ясно. Более того, пока такая перспектива кажется маловероятной. Обсуждение проблем, связанных с вторжением в нашу родословную родов и видов ардипитека (Ardipithecus ramidus), оррорина (Orrorin tugenensis), а также сахельантропа (Sahelanthropus tchadensis) только начинается, и пока ещё нуждается в дополнительном подтверждении даже сам тезис о том, что эти новые таксоны действительно относятся к гоминидам. Вывод о небольшой относительной величине зубов у орроринов тоже подлежит дальнейшей проверке, поскольку он прямо зависит от точности реконструкции размеров тела, а подобные реконструкции всегда небесспорны. Наконец, и увеличение возраста семейства гоминид до 6–7 млн лет, в принципе допустимое, тем не менее также сопряжено с некоторыми проблемами. Принимая его, придётся признать, что генетические датировки момента расхождения эволюционных путей предков человека и предков шимпанзе (см. табл. 2) в большинстве своём неверны и занижают возраст этого события. Таким образом, ни в коем случае не отказывая с порога представителям родов ардипитек (Ardipithecus), оррорин (Orrorin) и сахельантроп (Sahelanthropus) в праве «баллотироваться» в наши предки, не стоит вместе с тем и торопиться свергать австралопитеков с давно обжитого ими пьедестала, тем более что заменить их в этом случае будет просто некем.
Читайте также
Приложение 3. ВСЕОБЩАЯ ДЕКЛАРАЦИЯ О ГЕНОМЕ ЧЕЛОВЕКА И ПРАВАХ ЧЕЛОВЕКА
Приложение 3. ВСЕОБЩАЯ ДЕКЛАРАЦИЯ О ГЕНОМЕ ЧЕЛОВЕКА И ПРАВАХ ЧЕЛОВЕКА 3 декабря 1997 г.ВСЕОБЩАЯ ДЕКЛАРАЦИЯ О ГЕНОМЕ ЧЕЛОВЕКА И ПРАВАХ ЧЕЛОВЕКАГенеральная конференция,напоминая, что в преамбуле Устава ЮНЕСКО провозглашаются «демократические принципы уважения достоинства
Приложение 3. ВСЕОБЩАЯ ДЕКЛАРАЦИЯ О ГЕНОМЕ ЧЕЛОВЕКА И ПРАВАХ ЧЕЛОВЕКА
Приложение 3. ВСЕОБЩАЯ ДЕКЛАРАЦИЯ О ГЕНОМЕ ЧЕЛОВЕКА И ПРАВАХ ЧЕЛОВЕКА 3 декабря 1997 г.ВСЕОБЩАЯ ДЕКЛАРАЦИЯ О ГЕНОМЕ ЧЕЛОВЕКА И ПРАВАХ ЧЕЛОВЕКА Генеральная конференция,напоминая, что в преамбуле Устава ЮНЕСКО провозглашаются «демократические принципы уважения достоинства
Крупная дичь (буйволы и антилопы). Странные существа под упавшими стволами. Малые представители кошачьего рода
Крупная дичь (буйволы и антилопы). Странные существа под упавшими стволами. Малые представители кошачьего рода Сумеете ли вы провести трехтонный танк через любой лес так, чтобы ни одна веточка не хрустнула? Думаю, что это вам не удастся ни при каких обстоятельствах. Но
САМЫЕ РЕДКИЕ ПРЕДСТАВИТЕЛИ СЕМЕЙСТВА КОШАЧЬИХ
САМЫЕ РЕДКИЕ ПРЕДСТАВИТЕЛИ СЕМЕЙСТВА КОШАЧЬИХ 1 Амурский леопард2 Анталийский леопард3 Азиатский гепард4 Восточная пума5 Флоридская пума6 Северо-африканский леопард7 Сибирский тигр8 Южно-арабский леопард9 Южно-китайский тигр1 Суматрский
Первые обезьяны
Первые обезьяны В раннем эоцене (54–45 млн лет назад) в пределах отряда приматов выделяется уже множество семейств, родов и видов, среди которых есть и предки современных лемуров и долгопятов. Обычно этих ранних полуобезьян делят на лемуриформных (лемуры и их предки) и
Первые европейцы
Первые европейцы В раннем палеолите[36] Европа никак не могла претендовать на статус центра цивилизации. Она была скорее задворками Ойкумены, «медвежьим углом». Но все же и сюда добирались отголоски прогресса, ковавшегося в те времена в основном в Африке.Древнейшим
Как давно появились первые прямоходящие обезьяны – самые ранние предшественники человека?
Как давно появились первые прямоходящие обезьяны – самые ранние предшественники человека? Первой прямоходящей обезьяной на нашей планете был ореопитек – вид вымерших высших приматов. Известен он с 1872 года, когда горняки из провинции Тоскана (Италия) извлекли на
Первые опыты
Первые опыты И вот — нашлась «искра».Ею оказалась работа врача Леона Дюфура, увлекавшегося энтомологией и старательно изучавшего анатомию насекомых и пауков, их образ жизни, развитие и повадки. В этой работе Дюфур описывал свои наблюдения над осой-церцерис, охотящейся
Первые исследования ГМО
Первые исследования ГМО Вопрос биотехнологий и генетически модифицированных растений и других форм жизни впервые возник в ходе лабораторных исследований в США в начале 1970-х годов. В течение 1980-х администрация Рейгана проводила экономическую политику, практически
Первые признаки
Таблица 7. Гены, принимающие участие в образовании и функционировании ряда клеток, тканей и органов человека (по данным проекта «Геном человека» на 2000 г.)
Таблица 7. Гены, принимающие участие в образовании и функционировании ряда клеток, тканей и органов человека (по данным проекта «Геном человека» на
Первые неудачи
Первые неудачи Стремление все увидеть своими глазами побуждает Вишневского с командировкой университета отправиться за границу. В своем отчете, представленном по возвращении, он пишет:«Отправляясь в поездку, я ставил своей задачей знакомство с теми отделами
Первые аквариумы
Первые аквариумы Как-то в мае, когда я прогуливался в районе рыбных прудов у речушки Ракош, один мальчуган выудил маленькую — в пять сантиметров — щучку. Я тотчас выкупил у него рыбешку за форинт. В тот же день я оборудовал пятилитровый аквариум для щуки и двух солнечных
Первые результаты
Первые результаты За два года до сканирования Ричи мы разработали задачи, с помощью которых хотели изучить дисфункцию мозга у психопатов. В частности, мы хотели создать такие задачи, которые бы задействовали лимбическую систему мозга, которая в основном связана



